This is a modern-English version of Response in the Living and Non-Living, originally written by Bose, Jagadis Chandra.
It has been thoroughly updated, including changes to sentence structure, words, spelling,
and grammar—to ensure clarity for contemporary readers, while preserving the original spirit and nuance. If
you click on a paragraph, you will see the original text that we modified, and you can toggle between the two versions.
Scroll to the bottom of this page and you will find a free ePUB download link for this book.
Transcriber’s note: Four likely printer errors have been corrected; these are on pages 46, 115, 176 and 186, marked like this. The inconsistent hyphenation of “break-down”, “electro-motive” and “vibration-head” is as in the original. Some of the illustrations had to be moved up or down a few paragraphs from their position in the original; the hyperlinked page numbers in the List of Illustrations point to the original locations, but the hyperlinked figure numbers point to where the figures are now.
Transcriber’s note: Four likely printer errors have been corrected; these are on pages 46, 115, 176 and 186, marked like this. The inconsistent hyphenation of “break-down,” “electro-motive,” and “vibration-head” is as in the original. Some of the illustrations had to be moved up or down a few paragraphs from their position in the original; the hyperlinked page numbers in the List of Illustrations point to the original locations, but the hyperlinked figure numbers point to where the figures are now.
RESPONSE IN THE LIVING
AND NON-LIVING
BY
JAGADIS CHUNDER BOSE, M.A.(Cantab.), D.Sc.(London.)
PROFESSOR, PRESIDENCY COLLEGE, CALCUTTA
WITH ILLUSTRATIONS
WITH ILLUSTRATIONS
LONGMANS, GREEN, AND CO.
39 PATERNOSTER ROW, LONDON
NEW YORK AND BOMBAY
1902
All rights reserved
LONGMANS, GREEN, AND CO..
39 Paternoster Row, London
NEW YORK AND BOMBAY
1902
All rights reserved
‘The real is one: wise men call it variously’
'The real is one: wise people describe it in different ways'
Rig Veda
Rig Veda
To my Countrymen
This Work is Dedicated
To my fellow countrymen
This work is dedicated
PREFACE
I have in the present work put in a connected and a more complete form results, some of which have been published in the following Papers:
I have compiled a connected and more complete version of the results in this work, some of which have been published in the following papers:
- ‘De la Généralité des Phénomènes Moléculaires produits par l’Electricité sur la matière Inorganique et sur la matière Vivante.’ (Travaux du Congrès International de Physique. Paris, 1900.)
- ‘On the Similarity of Effect of Electrical Stimulus on Inorganic and Living Substances.’ (Report, Bradford Meeting British Association, 1900.—Electrician.)
- ‘Response of Inorganic Matter to Stimulus.’ (Friday Evening Discourse, Royal Institution, May 1901.)
- ‘On Electric Response of Inorganic Substances. Preliminary Notice.’ (Royal Society, June 1901.)
- ‘On Electric Response of Ordinary Plants under Mechanical Stimulus.’ (Journal Linnean Society, 1902.)
- ‘Sur la Réponse Electrique dans les Métaux, les Tissus Animaux et Végétaux.’ (Société de Physique, Paris, 1902.)
- ‘On the Electro-Motive Wave accompanying Mechanical Disturbance in Metals in contact with Electrolyte.’ (Proceedings Royal Society, vol. 70.)
- ‘On the Strain Theory of Vision and of Photographic Action.’ (Journal Royal Photographic Society, vol. xxvi.)
J. C. Bose.
J.C. Bose.
Davy-Faraday Laboratory, Royal Institution,
London: May 1902.
Davy-Faraday Lab, Royal Institution,
London: May 1902.
CONTENTS
CHAPTER I
| PAGE | |
Mechanical response—Different kinds of stimuli—Myograph—Characteristics of response-curve: period, amplitude, form—Modification of response-curves Mechanical response—Different types of stimuli—Myograph—Characteristics of response curve: period, amplitude, shape—Modification of response curves | 1 | |
CHAPTER II
| ||
Conditions for obtaining electric response—Method of injury—Current of injury—Injured end, cuproid: uninjured, zincoid—Current of response in nerve from more excited to less excited—Difficulties of present nomenclature—Electric recorder—Two types of response, positive and negative—Universal applicability of electric mode of response—Electric response a measure of physiological activity—Electric response in plants Conditions for getting an electric response—Method of injury—Injury current—Injured end, cupric: uninjured, zinc—Response current in the nerve from more excited to less excited—Challenges with current terminology—Electric recorder—Two types of responses, positive and negative—Universal applicability of electric response—Electric response as an indicator of physiological activity—Electric response in plants. | 5 | |
CHAPTER III
| ||
Negative variation—Response recorder—Photographic recorder—Compensator—Means of graduating intensity of stimulus—Spring-tapper and torsional vibrator—Intensity of stimulus dependent on amplitude of vibration—Effectiveness of stimulus dependent on rapidity also Negative variation—Response recorder—Photographic recorder—Compensator—Ways to adjust stimulus intensity—Spring-tapper and torsional vibrator—Stimulus intensity depends on vibration amplitude—Stimulus effectiveness also depends on speed. | 17 | |
CHAPTER IV
| ||
Method of block—Advantages of block method—Plant response a physiological phenomenon—Abolition of response by anæsthetics and poisons—Abolition of response when plant is killed by hot water Method of block—Advantages of block method—Plant response is a physiological phenomenon—Response is eliminated by anesthetics and poisons—Response is abolished when the plant is killed by hot water | 27 | |
CHAPTER V
| ||
Effect of single stimulus—Superposition of stimuli—Additive effect—Staircase effect—Fatigue—No fatigue when sufficient interval between stimuli—Apparent fatigue when stimulation frequency is increased—Fatigue under continuous stimulation Effect of a single stimulus—Layering of stimuli—Cumulative effect—Staircase effect—Tiredness—No tiredness with a sufficient break between stimuli—Seeming tiredness when the stimulation frequency goes up—Tiredness with continuous stimulation | 35 | |
CHAPTER VI
| ||
Diphasic variation—Positive after-effect and positive response—Radial E.M. variation Diphasic variation—Positive after-effect and positive response—Radial E.M. variation | 44 | |
CHAPTER VII
| ||
Increased response with increasing stimulus—Apparent diminution of response with excessively strong stimulus Increased response with stronger stimuli—Noticeable decrease in response with overly strong stimuli | 51 | |
CHAPTER VIII
| ||
Effect of very low temperature—Influence of high temperature—Determination of death-point—Increased response as after-effect of temperature variation—Death of plant and abolition of response by the action of steam Effect of very low temperature—Influence of high temperature—Determining the death point—Increased response as an after-effect of temperature change—Death of the plant and loss of response due to steam action | 59 | |
CHAPTER IX
| ||
Effect of anæsthetics, a test of vital character of response—Effect of chloroform—Effect of chloral—Effect of formalin—Method in which response is unaffected by variation of resistance—Advantage of block method—Effect of dose Effect of anesthetics, a test of vital character of response—Effect of chloroform—Effect of chloral—Effect of formalin—Method in which response is unaffected by variation of resistance—Advantage of block method—Effect of dose | 71 | |
CHAPTER X
| ||
Is response found in inorganic substances?—Experiment on tin, block method—Anomalies of existing terminology—Response by method of depression—Response by method of exaltation Is response found in inorganic substances?—Experiment on tin, block method—Anomalies of existing terminology—Response by method of depression—Response by method of exaltation | 81 | |
CHAPTER XI
| ||
Conditions of obtaining quantitative measurements—Modification of the block method—Vibration cell—Application of stimulus—Graduation of the intensity of stimulus—Considerations showing that electric response is due to molecular disturbance—Test experiment—Molecular voltaic cell Conditions for obtaining quantitative measurements—Modification of the block method—Vibration cell—Application of stimulus—Grading the intensity of stimulus—Factors indicating that electric response is caused by molecular disturbance—Test experiment—Molecular voltaic cell | 91 | |
CHAPTER XII
| ||
Preparation of wire—Effect of single stimulus Preparation of wire—Effect of a single stimulus | 100 | |
CHAPTER XIII
| ||
Effects of molecular inertia—Prolongation of period of recovery by overstrain—Molecular model—Reduction of molecular sluggishness attended by quickened recovery and heightened response—Effect of temperature—Modification of latent period and period of recovery by the action of chemical reagents—Diphasic variation Effects of molecular inertia—Longer recovery time due to overstrain—Molecular model—Decreased molecular sluggishness leads to faster recovery and improved response—Impact of temperature—Changes in the latent period and recovery time caused by chemical agents—Diphasic variation | 104 | |
CHAPTER XIV
| ||
Fatigue in metals—Fatigue under continuous stimulation—Staircase effect—Reversed responses due to molecular modification in nerve and in metal, and their transformation into normal after continuous stimulation—Increased response after continuous stimulation Fatigue in metals—Fatigue from constant stimulation—Staircase effect—Opposite reactions caused by molecular changes in nerves and metals, and their return to normal after ongoing stimulation—Heightened response after constant stimulation | 118 | |
CHAPTER XV
| ||
Relation between stimulus and response—Magnetic analogue—Increase of response with increasing stimulus—Threshold of response—Superposition of stimuli—Hysteresis Relation between stimulus and response—Magnetic analogy—Increase in response with increased stimulus—Response threshold—Superimposing stimuli—Hysteresis | 131 | |
CHAPTER XVI
| ||
Action of chemical reagents—Action of stimulants on metals—Action of depressants on metals—Effect of ‘poisons’ on metals—Opposite effect of large and small doses Action of chemical reagents—Action of stimulants on metals—Action of depressants on metals—Effect of ‘poisons’ on metals—Opposite effect of large and small doses | 139 | |
CHAPTER XVII
| ||
Visual impulse: (1) chemical theory; (2) electrical theory—Retinal currents—Normal response positive—Inorganic response under stimulus of light—Typical experiment on the electrical effect induced by light Visual impulse: (1) chemical theory; (2) electrical theory—Retinal currents—Normal response is positive—Inorganic response triggered by light—Typical experiment on the electrical effect caused by light. | 148 | |
CHAPTER XVIII
| ||
Effect of temperature—Effect of increasing length of exposure—Relation between intensity of light and magnitude of response—After-oscillation—Abnormal effects: (1) preliminary negative twitch; (2) reversal of response; (3) transient positive twitch on cessation of light; (4) decline and reversal—Résumé Effect of temperature—Effect of longer exposure time—Relationship between light intensity and level of response—After-oscillation—Unusual effects: (1) initial negative twitch; (2) response reversal; (3) temporary positive twitch when light stops; (4) decline and reversal—Summary | 158 | |
CHAPTER XIX
| ||
Effect of light of short duration—After-oscillation—Positive and negative after-images—Binocular alternation of vision—Period of alternation modified by physical condition—After-images and their revival—Unconscious visual impression. Effect of short-duration light—After-oscillation—Positive and negative after-images—Binocular vision alternation—Duration of alternation affected by physical condition—After-images and their revival—Unconscious visual impression. | 170 | |
CHAPTER XX
| 181 | |
INDEX | 193 |
ILLUSTRATIONS
| FIG. | PAGE | |
|---|---|---|
| 1. | Mechanical Lever Recorder | 3 |
| 2. | Electric Method of Detecting Nerve Response | 6 |
| 3. | Diagram showing Injured End of Nerve Corresponds to Copper in a Voltaic Element | 8 |
| 4. | Electric Recorder | 11 |
| 5. | Simultaneous Record of Mechanical and Electrical Responses | 13 |
| 6. | Negative Variation in Plants | 19 |
| 7. | Photographic Record of Negative Variation in Plants | 20 |
| 8. | Response Recorder | 21 |
| 9. | The Compensator | 22 |
| 10. | The Spring-tapper | 23 |
| 11. | The Torsional Vibrator | 24 |
| 12. | Response in Plant to Mechanical Tap or Vibration | 25 |
| 13. | Influence of Suddenness on the Efficiency of Stimulus | 26 |
| 14. | The Method of Block | 28 |
| 15. | Response in Plant completely Immersed under Water | 29 |
| 16. | Uniform Responses in Plant | 36 |
| 17. | Fusion of Effect under Rapidly Succeeding Stimuli in Muscle and in Plant | 36 |
| 18. | Additive Effect of Singly Ineffective Stimuli on Plant | 37 |
| 19. | ‘Staircase Effect’ in Plant | 37 |
| 20. | Appearance of Fatigue in Plant under Shortened Period of Rest | 39 |
| 21. | Fatigue in Celery | 40 |
| 22. | Fatigue in Cauliflower-stalk | 41 |
| 23. | Fatigue from Previous Overstrain | 41 |
| 24. | Fatigue under Continuous Stimulation in Celery | 42 |
| 25. | Effect of Rest in Removal of Fatigue in Plant | 43 |
| 26. | Diphasic Variation in Plant | 46 |
| 27, 28. | Abnormal Positive Responses in Stale Plant transformed into Normal Negative Under Strong Stimulation | 48, 49 |
| 29. | Radial E.M. Variation | 50 |
| 30. | Curves showing the Relation between Intensity of Stimulus and Response in Muscle and Nerve | 52 |
| 31. | Increasing Responses to Increasing Stimuli (Taps) in Plants | 52 |
| 32. | Increasing Responses to Increasing Vibrational Stimuli in Plants | 53 |
| 33. | Responses to Increasing Stimuli in Fresh and Stale Specimens of Plants | 54 |
| 34. | Apparent Diminution of Response caused by Fatigue under Strong Stimulation | 57 |
| 35. | Diminution of Response in Eucharis Lily at Low Temperature | 61 |
| 36. | Records showing the Difference in the Effects of Low Temperature on Ivy, Holly, and Eucharis Lily | 62 |
| 37. | Plant Chamber for Studying the Effect of Temperature and Anæsthetics | 64 |
| 38. | Effect of High Temperature on Plant Response | 64 |
| 39. | After-effect on the Response due to Temperature Variation | 66 |
| 40. | Records of Responses in Eucharis Lily during Rise and Fall of Temperature | 67 |
| 41. | Curve showing Variation of Sensitiveness during a Cycle of Temperature Variation | 68 |
| 42. | Record of Effect of Steam in Abolition of Response at Death of Plant | 69 |
| 43. | Effect of Chloroform on Nerve Response | 72 |
| 44. | Effect of Chloroform on the Responses of Carrot | 74 |
| 45. | Action of Chloral Hydrate on Plant Responses | 75 |
| 46. | Action of Formalin on Radish | 75 |
| 47. | Action of Sodium Hydrate in Abolishing the Response in Plant | 78 |
| 48. | Stimulating Action of Poison in Small Doses in Plants | 79 |
| 49. | The Poisonous Effect of Stronger Dose of KOH | 79 |
| 50. | Block Method for obtaining Response in Tin | 83 |
| 51. | Response To Mechanical Stimulation in a Zn-Cu Couple | 85 |
| 52. | Electric Response in Metal by the Method of Relative Depression (Negative Variation) | 88 |
| 53. | Method of Relative Exaltation | 89 |
| 54. | Various Cases of Positive and Negative Variation | 90 |
| 55. | Modifications of the Block Method for Exhibiting Electric Response in Metals | 93 |
| 56. | Equal and Opposite Responses given by Two Ends of the Wire | 95 |
| 57. | Top View of the Vibration Cell | 96 |
| 58. | Influence of Annealing in the Enhancement of Response in Metals | 101 |
| 59. | Uniform Electric Responses in Metals | 102 |
| 60. | Persistence of After-effect | 105 |
| 61. | Prolongation of Period of Recovery after Overstrain | 106 |
| 62. | Molecular Model | 107 |
| 63, 64. | Effects of Removal of Molecular Sluggishness in Quickened Recovery and Heightened Response in Metals | 109, 110 |
| 65. | Effect of Temperature on Response in Metals | 111 |
| 66. | Diphasic Variation in Metals | 113 |
| 67. | Negative, Diphasic, and Positive Resultant Response in Metals | 115 |
| 68. | Continuous Transformation from Negative to Positive through Intermediate Diphasic Response | 116 |
| 69. | Fatigue in Muscle | 118 |
| 70. | Fatigue in Platinum | 118 |
| 71. | Fatigue in Tin | 119 |
| 72. | Appearance of Fatigue due to Shortening the Period of Recovery | 120 |
| 73. | Fatigue in Metal under Continuous Stimulation | 121 |
| 74. | ‘Staircase’ Response in Muscle and in Metal | 122 |
| 75. | Abnormal Response in Nerve converted into Normal under Continued Stimulation | 124 |
| 76, 77. | Abnormal Response in Tin and Platinum converted into Normal under Continued Stimulation | 125 |
| 78. | Gradual Transition from Abnormal to Normal Response in Platinum | 126 |
| 79. | Increase of Response in Nerve after Continuous Stimulation | 127 |
| 80, 81. | Response in Tin and Platinum Enhanced after Continuous Stimulation | 127, 128 |
| 82. | Magnetic Analogue | 132 |
| 83, 84. | Records of Responses to Increasing Stimuli in Tin | 134, 135 |
| 85. | Ineffective Stimulus becoming Effective by Superposition | 135 |
| 86. | Incomplete and Complete Fusion of Effects | 136 |
| 87. | Cyclic Curve for Maximum Effects showing Hysteresis | 137 |
| 88. | Action of Poison in Abolishing Response in Nerve | 139 |
| 89. | Action of Stimulant on Tin | 141 |
| 90. | Action of Stimulant on Platinum | 142 |
| 91. | Depressing Effect of KBr on Tin | 143 |
| 92. | Abolition of Response in Metals by ‘Poison’ | 143 |
| 93. | ‘Molecular Arrest’ by the Action of ‘Poison’ | 145 |
| 94. | Opposite Effects of Small and Large Doses on the Response in Metals | 146 |
| 95. | Retinal Response to Light | 150 |
| 96. | Response of Sensitive Cell to Light | 152 |
| 97. | Typical Experiment on the E.M. Variation Produced by Light | 154 |
| 98. | Modification of the Photo-sensitive Cell | 155 |
| 99. | Responses in Frog’s Retina | 156 |
| 100. | Responses in Sensitive Photo-cell | 157 |
| 101. | Effect of Temperature on the Response to Light Stimulus | 159 |
| 102. | Effect of Duration of Exposure on the Response | 159 |
| 103. | Responses of Sensitive Cell to Increasing Intensities of Light | 161 |
| 104. | Relation between the Intensity of Light And Magnitude of Response | 162 |
| 105. | After-oscillation | 163 |
| 106. | Transient Positive Increase of Response in the Frog’s Retina on the Cessation of Light | 164 |
| 107. | Transient Positive Increase of Response in the Sensitive Cell | 165 |
| 108. | Decline under the Continuous Action of Light | 166 |
| 109. | Certain After-effects of Light | 168 |
| 110. | After-effect of Light of Short Duration | 172 |
| 111. | Stereoscopic Design for the Exhibition of Binocular Alternation of Vision | 176 |
| 112. | Uniform Responses in Nerve, Plant, and Metal | 184 |
| 113. | Fatigue in Muscle, Plant, and Metal | 185 |
| 114. | ‘Staircase’ Effect in Muscle, Plant, and Metal | 186 |
| 115. | Increase of Response after Continuous Stimulation in Nerve and Metal | 186 |
| 116. | Modified Abnormal Response in Nerve and Metal Transformed into Normal Response after Continuous Stimulation | 187 |
| 117. | Action of the same ‘Poison’ in the Abolition of Response in Nerve, Plant, and Metal | 189 |
RESPONSE
IN THE
LIVING AND NON-LIVING
CHAPTER I
THE MECHANICAL RESPONSE OF LIVING SUBSTANCES
- Mechanical response
- —Different kinds of stimuli
- —Myograph
- —Characteristics of response-curve: period, amplitude, form
- —Modification of response-curves.
One of the most striking effects of external disturbance on certain types of living substance is a visible change of form. Thus, a piece of muscle when pinched contracts. The external disturbance which produced this change is called the stimulus. The body which is thus capable of responding is said to be irritable or excitable. A stimulus thus produces a state of excitability which may sometimes be expressed by change of form.
One of the most noticeable effects of external disturbance on certain types of living matter is a visible change in shape. For example, when a piece of muscle is pinched, it contracts. The external disturbance that causes this change is known as a stimulus. The body that can respond in this way is described as irritable or excitable. A stimulus, therefore, creates a state of excitability that can sometimes be shown through changes in form.
Mechanical response to different kinds of stimuli.—This reaction under stimulus is seen even in the lowest organisms; in some of the amœboid rhizopods, for instance. These lumpy protoplasmic bodies, usually elongated while creeping, if mechanically jarred, contract into a spherical form. If, instead of mechanical disturbance, we apply salt solution, they again contract, in the same way as before. Similar effects are produced by sudden illumination, or by rise of temperature, or by electric shock. A living substance may thus be put into an excitatory state by either mechanical, chemical, thermal, electrical, or light stimulus. Not only does the point stimulated show the effect of stimulus, but that effect may sometimes be conducted even to a considerable distance. This power of conducting stimulus, though common to all living substances, is present in very different degrees. While in some forms of animal tissue irritation spreads, at a very slow rate, only to points in close neighbourhood, in other forms, as for example in nerves, conduction is very rapid and reaches far.
Mechanical response to different kinds of stimuli.—This reaction to stimuli occurs even in the simplest organisms, like some of the amœboid rhizopods. These irregular protoplasmic bodies, which usually stretch out while moving, will contract into a spherical shape if they are mechanically disturbed. Similarly, if we apply a salt solution instead of a mechanical disturbance, they will contract in the same way. Sudden exposure to light, increasing temperature, or electric shock can produce similar effects. A living substance can be put into an excitatory state by mechanical, chemical, thermal, electrical, or light stimuli. Not only does the area that is stimulated show the effects of the stimulus, but this effect can sometimes be transmitted even over considerable distances. This ability to transmit stimuli, although present in all living substances, varies widely in degree. While in some types of animal tissue irritation spreads slowly only to nearby areas, in others, like nerves, the transmission is very fast and can reach far.
The visible mode of response by change of form may perhaps be best studied in a piece of muscle. When this is pinched, or an electrical shock is sent through it, it becomes shorter and broader. A responsive twitch is thus produced. The excitatory state then disappears, and the muscle is seen to relax into its normal form.
The visible way a muscle responds by changing its shape can be best observed in a piece of muscle tissue. When you pinch it or send an electrical shock through it, it gets shorter and wider. This creates a noticeable twitch. Once the excitement wears off, the muscle relaxes back into its normal shape.
Mechanical lever recorder.—In the case of contraction of muscle, the effect is very quick, the twitch takes place in too short a time for detailed observation by ordinary means. A myographic apparatus is therefore used, by means of which the changes in the muscle are self-recorded. Thus we obtain a history of its change and recovery from the change. The muscle is connected to one end of a writing lever. When the muscle contracts, the tracing point is pulled up in one direction, say to the right. The extent of this pull depends on the amount of contraction. A band of paper or a revolving drum-surface moves at a uniform speed at right angles to the direction of motion of the writing lever. When the muscle recovers from the stimulus, it relaxes into its original form, and the writing point traces the recovery as it moves now to the left, regaining its first position. A curve is thus described, the rising portion of which is due to contraction, and the falling portion to relaxation or recovery. The ordinate of the curve represents the intensity of response, and the abscissa the time (fig. 1).
Mechanical lever recorder.—When a muscle contracts, the effect happens very quickly, and the twitch occurs in such a short time that ordinary observation methods can’t capture it in detail. Therefore, a myographic device is used to self-record changes in the muscle. This gives us a record of its change and recovery. One end of a writing lever is connected to the muscle. When the muscle contracts, the tracing point moves upward in one direction, like to the right. The height of this pull is determined by how much the muscle contracts. A strip of paper or a rotating drum surface moves at a consistent speed perpendicular to the writing lever's movement. When the muscle relaxes after the stimulus, it returns to its original shape, and the writing point traces this recovery as it moves back to the left, returning to its initial position. This creates a curve, with the rising part showing contraction and the falling part showing relaxation or recovery. The vertical axis of the curve indicates the intensity of the response, while the horizontal axis represents the time (fig. 1).

Fig. 1.—Mechanical Lever Recorder
Fig. 1.—Mechanical Lever Recorder
The muscle M with the attached bone is securely held at one end, the other end being connected with the writing lever. Under the action of stimulus the contracting muscle pulls the lever and moves the tracing point to the right over the travelling recording surface P. When the muscle recovers from contraction, the tracing point returns to its original position. See on P the record of muscle curve.
The muscle M, attached to a bone at one end, is firmly secured, while the other end connects to the writing lever. When stimulated, the contracting muscle pulls the lever, moving the tracing point to the right across the travelling recording surface P. Once the muscle relaxes, the tracing point goes back to its original position. See the record of the muscle curve on P.
Characteristics of the response-curve: (1) Period, (2) Amplitude, (3) Form.—Just as a wave of sound is characterised by its (1) period, (2) amplitude, and (3) form, so may these response-curves be distinguished from each other. As regards the period, there is an enormous variation, corresponding to the functional activity of the muscle. For instance, in tortoise it may be as high as a second, whereas in the wing-muscles of many insects it is as small as 1/300 part of a second. ‘It is probable that a continuous graduated scale might, as suggested by Hermann, be drawn up in the animal kingdom, from the excessively rapid contraction of insects to those of tortoises and hibernating dormice.’[1] Differences in form and amplitude of curve are well illustrated by various muscles of the tortoise. The curve for the muscle of the neck, used for rapid withdrawal of the head on approach of danger, is quite different from that of the pectoral muscle of the same animal, used for its sluggish movements.
Characteristics of the response-curve: (1) Period, (2) Amplitude, (3) Form.—Just like a sound wave is defined by its (1) period, (2) amplitude, and (3) shape, these response curves can also be differentiated from one another. When it comes to the period, there is a huge range that reflects the muscle's functional activity. For example, in a tortoise, it can be as long as a second, while in the wing muscles of many insects, it can be as short as 1/300 of a second. ‘It’s likely that a continuous graduated scale could, as Hermann suggested, be created in the animal kingdom, ranging from the extremely rapid contractions of insects to those of tortoises and hibernating dormice.’[1] Differences in the shape and amplitude of the curve are well demonstrated by different muscles in the tortoise. The curve for the neck muscle, used for quickly withdrawing the head in response to danger, is quite different from that of the pectoral muscle, which is used for its slow movements.
Again, progressive changes in the same muscle are well seen in the modifications of form which consecutive muscle-curves gradually undergo. In a dying muscle, for example, the amplitude of succeeding curves is continuously diminished, and the curves themselves are elongated. Numerous illustrations will be seen later, of the effect, in changing the form of the curve, of the increased excitation or depression produced by various agencies.
Again, gradual changes in the same muscle are clearly visible in the changes in shape that the muscle curves go through over time. In a dying muscle, for instance, the size of the following curves steadily decreases, while the curves themselves become longer. Many examples will be shown later, demonstrating how changes in the curve's shape are influenced by increased stimulation or suppression from different factors.
Thus these response records give us a means of studying the effect of stimulus, and the modification of response, under varying external conditions, advantage being taken of the mechanical contraction produced in the tissue by the stimulus. But there are other kinds of tissue where the excitation produced by stimulus is not exhibited in a visible form. In order to study these we have to use an altogether independent method, the method of electric response.
Thus, these response records provide a way for us to study the effects of stimuli and how responses change under different external conditions, taking advantage of the mechanical contraction caused in the tissue by the stimulus. However, there are other types of tissue where the excitement generated by the stimulus isn't shown visibly. To study these, we need to use a completely different method, the method of electric response.
CHAPTER II
ELECTRIC RESPONSE
- Conditions for obtaining electric response
- —Method of injury
- —Current of injury
- —Injured end, cuproid: uninjured, zincoid
- —Current of response in nerve from more excited to less excited
- —Difficulties of present nomenclature
- —Electric recorder
- —Two types of response, positive and negative
- —Universal applicability of electric mode of response
- —Electric response a measure of physiological activity
- —Electric response in plants.
Unlike muscle, a length of nerve, when mechanically or electrically excited, does not undergo any visible change. That it is thrown into an excitatory state, and that it conducts the excitatory disturbance, is shown however by the contraction produced in an attached piece of muscle, which serves as an indicator.
Unlike muscle, a section of nerve doesn’t show any visible change when it's stimulated mechanically or electrically. However, the fact that it enters an excitatory state and carries the excitatory signal is evident from the contraction that occurs in a connected piece of muscle, which acts as an indicator.
But the excitatory effect produced in the nerve by stimulus can also be detected by an electrical method. If an isolated piece of nerve be taken and two contacts be made on its surface by means of non-polarisable electrodes at A and B, connection being made with a galvanometer, no current will be observed, as both A and B are in the same physico-chemical condition. The two points, that is to say, are iso-electric.
But the excitatory effect created in the nerve by a stimulus can also be detected using an electrical method. If you take a piece of isolated nerve and make two contacts on its surface with non-polarizable electrodes at A and B, connecting it to a galvanometer, you won’t see any current, since both A and B are in the same physico-chemical condition. In other words, the two points are iso-electric.
If now the nerve be excited by stimulus, similar disturbances will be evoked at both A and B. If, further, these disturbances, reaching A and B almost simultaneously, cause any electrical change, then, similar changes taking place at both points, and there being thus no relative difference between the two, the galvanometer will still indicate no current. This null-effect is due to the balancing action of B as against A. (See fig. 2, a.)
If the nerve is stimulated now, similar disturbances will occur at both A and B. If these disturbances reach A and B almost at the same time and cause any electrical change, then, similar changes happening at both points mean there will be no relative difference between the two, so the galvanometer will still show no current. This null effect is due to the balancing action of B against A. (See fig. 2, a.)
Conditions for obtaining electric response.—If then we wish to detect the response by means of the galvanometer, one means of doing so will lie in the abolition of this balance, which may be accomplished by making one of the two points, say B, more or less permanently irresponsive. In that case, stimulus will cause greater electrical disturbance at the more responsive point, say A, and this will be shown by the galvanometer as a current of response. To make B less responsive we may injure it by means of a cross-sectional cut, a burn, or the action of strong chemical reagents.
Conditions for obtaining electric response.—If we want to detect the response using a galvanometer, one way to do this is by eliminating the balance. This can be achieved by making one of the two points, say B, somewhat permanently unresponsive. In this case, a stimulus will create a larger electrical disturbance at the more responsive point, say A, which will be indicated by the galvanometer as a current of response. To make B less responsive, we might damage it by making a cross-sectional cut, applying a burn, or using strong chemical reagents.

Fig. 2.—Electric Method of Detecting Nerve Response
Fig. 2.—Electric Method of Detecting Nerve Response
(a) Iso-electric contacts; no current in the galvanometer.
(b) The end B injured; current of injury from B to A: stimulation gives rise to
an action current from A to B.
(c) Non-polarisable electrode.
(a) Iso-electric contacts; no current in the galvanometer.
(b) The end B is damaged; the injury current flows from B to A: stimulation causes an action current to flow from A to B.
(c) Non-polarizable electrode.
Current of injury.—We shall revert to the subject of electric response; meanwhile it is necessary to say a few words regarding the electric disturbance caused by the injury itself. Since the physico-chemical conditions of the uninjured A and the injured B are now no longer the same, it follows that their electric conditions have also become different. They are no longer iso-electric. There is thus a more or less permanent or resting difference of electric potential between them. A current—the current of injury—is found to flow in the nerve, from the injured to the uninjured, and in the galvanometer, through the electrolytic contacts from the uninjured to the injured. As long as there is no further disturbance this current of injury remains approximately constant, and is therefore sometimes known as ‘the current of rest’ (fig. 2, b).
Current of injury.—We will return to the topic of electric response; for now, it’s important to mention a few things about the electric disruption caused by the injury itself. Since the physical and chemical conditions of the uninjured A and the injured B are no longer the same, it follows that their electric conditions have also changed. They are no longer iso-electric. This results in a more or less permanent or resting difference in electric potential between them. A current—the current of injury—flows in the nerve, from the injured to the uninjured, and in the galvanometer, through the electrolytic contacts from the uninjured to the injured. As long as there is no further disruption, this current of injury remains roughly constant, and is sometimes referred to as ‘the current of rest’ (fig. 2, b).
A piece of living tissue, unequally injured at the two ends, is thus seen to act like a voltaic element, comparable to a copper and zinc couple. As some confusion has arisen, on the question of whether the injured end is like the zinc or copper in such a combination, it will perhaps be well to enter upon this subject in detail.
A section of living tissue, injured unevenly at both ends, acts like a battery, similar to a copper and zinc pair. Since there has been some confusion about whether the injured end resembles the zinc or the copper in this setup, it might be helpful to discuss this topic in detail.
If we take two rods, of zinc and copper respectively, in metallic contact, and further, if the points A and B are connected by a strip of cloth s moistened with salt solution, it will be seen that we have a complete voltaic element. A current will now flow from B to A in the metal (fig. 3, a) and from A to B through the electrolyte s. Or instead of connecting A and B by a single strip of cloth s, we may connect them by two strips s s′, leading to non-polarisable electrodes E E′. The current will then be found just the same as before, i.e. from B to A in the metallic part, and from A through s s′ to B, the wire W being interposed, as it were, in the electrolytic part of the circuit. If now a galvanometer be interposed at O, the current will flow from B to A through the galvanometer, i.e. from right to left. But if we interpose the galvanometer in the electrolytic part of the circuit, that is to say, at W, the same current will appear to flow in the opposite direction. In fig. 3, c, the galvanometer is so interposed, and in this case it is to be noticed that when the current in the galvanometer flows from left to right, the metal connected to the left is zinc.
If we take two rods, one of zinc and the other of copper, and put them in metal contact, and then connect points A and B with a strip of cloth s soaked in salt solution, we create a complete voltaic element. A current will flow from B to A in the metal (fig. 3, a) and from A to B through the electrolyte s. Alternatively, instead of using just one strip of cloth s to connect A and B, we can use two strips s s′, which connect to non-polarizable electrodes E E′. The current will still flow the same way as before, meaning from B to A in the metallic part and from A through s s′ to B, with wire W acting as an intermediary in the electrolytic part of the circuit. If we place a galvanometer at O, the current will flow from B to A through the galvanometer, or from right to left. However, if we put the galvanometer in the electrolytic part of the circuit, specifically at W, the same current will seem to flow in the opposite direction. In fig. 3, c, the galvanometer is situated this way, and it's worth noting that when the current in the galvanometer flows from left to right, the metal on the left is zinc.
Compare fig. 3, d, where A B is a piece of nerve of which the B end is injured. The current in the galvanometer through the non-polarisable electrode is from left to right. The uninjured end is therefore comparable to the zinc in a voltaic cell (is zincoid), the injured being copper-like or cuproid.[2]
Compare fig. 3, d, where A B is a piece of nerve with the B end damaged. The current in the galvanometer flows from left to right through the non-polarizable electrode. The uninjured end is therefore similar to the zinc in a voltaic cell (is zincoid), while the injured end is comparable to copper or cuproid.[2]

Fig. 3.—Diagram showing the Correspondence between injured (B) and uninjured (A) contacts in Nerve, and Cu and Zn in a Voltaic Element
Fig. 3.—Diagram showing the relationship between injured (B) and uninjured (A) contacts in the nerve, and Cu and Zn in a voltaic element.
Comparison of (c) and (d) will show that the injured end of B in (d) corresponds with the Cu in (c).
Comparison of (c) and (d) will show that the injured end of B in (d) matches up with the Cu in (c).
If the electrical condition of, say, zinc in the voltaic couple (fig. 3, c) undergo any change (and I shall show later that this can be caused by molecular disturbance), then the existing difference of potential between A and B will also undergo variation. If for example the electrical condition of A approach that of B, the potential difference will undergo a diminution, and the current hitherto flowing in the circuit will, as a consequence, display a diminution, or negative variation.
If the electrical condition of, let's say, zinc in the voltaic couple (fig. 3, c) changes (and I’ll explain later that this can happen due to molecular disturbance), then the existing difference in potential between A and B will also change. For example, if the electrical condition of A gets closer to that of B, the potential difference will decrease, and the current that has been flowing in the circuit will, as a result, show a decrease or negative change.
Action current.—We have seen that a current of injury—sometimes known as ‘current of rest’—flows in a nerve from the injured to the uninjured, and that the injured B is then less excitable than the uninjured A. If now the nerve be excited, there being a greater effect produced at A, the existing difference of potential may thus be reduced, with a consequent diminution of the current of injury. During stimulation, therefore, a nerve exhibits a negative variation. We may express this in a different way by saying that a ‘current of action’ was produced in response to stimulus, and acted in an opposite direction to the current of injury (fig. 2, b). The action current in the nerve is from the relatively more excited to the relatively less excited.
Action current.—We've observed that an injury current—sometimes referred to as a ‘current of rest’—flows through a nerve from the injured area to the uninjured area, and that the injured B is less responsive than the uninjured A. If the nerve is stimulated, and a greater effect occurs at A, the existing potential difference can be reduced, which decreases the injury current. Therefore, during stimulation, a nerve shows a negative variation. We can also say that a ‘current of action’ is generated in response to the stimulus, moving in the opposite direction of the injury current (fig. 2, b). The action current in the nerve flows from the relatively more excited to the relatively less excited.
Difficulties of present nomenclature.—We shall deal later with a method by which a responsive current of action is obtained without any antecedent current of injury. ‘Negative variation’ has then no meaning. Or, again, a current of injury may sometimes undergo a change of direction (see note, p. 12). In view of these considerations it is necessary to have at our disposal other forms of expression by which the direction of the current of response can still be designated. Keeping in touch with the old phraseology, we might then call a current ‘negative’ that flowed from the more excited to the less excited. Or, bearing in mind the fact that an uninjured contact acts as the zinc in a voltaic couple, we might call it ‘zincoid,’ and the injured contact ‘cuproid.’ Stimulation of the uninjured end, approximating it to the condition of the injured, might then be said to induce a cuproid change.
Challenges with Current Terminology.—We'll discuss a method later that allows for a responsive action current to be generated without any prior injury current, making 'negative variation' irrelevant. Moreover, an injury current can sometimes reverse direction (see note, __A_TAG_PLACEHOLDER_0__). Given these points, we need new terms to indicate the direction of the response current. Sticking to older terminology, we might label a current as 'negative' if it flows from the more excited area to the less excited area. Also, since an uninjured contact acts like the zinc in a galvanic cell, we could call it 'zincoid,' while labeling the injured contact as 'cuproid.' Stimulating the uninjured end to mimic the injured condition could then be termed inducing a cuproid change.
The electric change produced in a normal nerve by stimulation may therefore be expressed by saying that there has been a negative variation, or that there was a current of action from the more excited to the less excited, or that stimulation has produced a cuproid change.
The electric change that occurs in a normal nerve when stimulated can be described as a negative variation, indicating an action current flowing from the more excited area to the less excited area, or it can be said that stimulation has caused a cuproid change.
The excitation, or molecular disturbance, produced by a stimulus has thus a concomitant electrical expres sion. As the excitatory state disappears with the return of the excitable tissue to its original condition, the current of action will gradually disappear.[3] The movement of the galvanometer needle during excitation of the tissue thus indicates a molecular upset by the stimulus; and the gradual creeping back of the galvanometer deflection exhibits a molecular recovery.
The excitement, or molecular disturbance, caused by a stimulus has an accompanying electrical expression. As the excitatory state fades and the excitable tissue returns to its original condition, the action current will slowly diminish. The movement of the galvanometer needle during the tissue excitation shows a molecular disruption from the stimulus, while the gradual return of the galvanometer deflection demonstrates a molecular recovery.
This transitory electrical variation constitutes the ‘response,’ and its intensity varies according to that of the stimulus.
This temporary electrical change is the 'response,' and its strength varies based on the stimulus.
Electric recorder.—We have thus a method of obtaining curves of response electrically. After all, it is not essentially very different from the mechanical method. In this case we use a magnetic lever (fig. 4, a), the needle of the galvanometer, which is deflected by the electromagnetic pull of the current, generated under the action of stimulus, just as the mechanical lever was deflected by the mechanical pull of the muscle contracting under stimulus.
Electric recorder.—So, we have a way to get response curves using electricity. Really, it's not that different from the mechanical method. Here, we use a magnetic lever (fig. 4, a), and the needle of the galvanometer is moved by the electromagnetic force of the current generated when a stimulus is applied, just like the mechanical lever was moved by the muscle's mechanical force when it contracted in response to a stimulus.
The accompanying diagram (fig. 4, b) shows how, under the action of stimulus, the current of rest undergoes a transitory diminution, and how on the cessation of stimulus there is gradual recovery of the tissue, as exhibited in the return of the galvanometer needle to its original position.
The accompanying diagram (fig. 4, b) shows how, when a stimulus is applied, the resting current decreases temporarily, and how after the stimulus stops, the tissue gradually recovers, which is seen in the galvanometer needle returning to its starting position.

Fig. 4.—Electric Recorder
Fig. 4.—Digital Recorder
(a) M muscle; A uninjured, B injured ends. E E′ non-polarising electrodes connecting A and B with galvanometer G. Stimulus produces ‘negative variation’ of current of rest. Index connected with galvanometer needle records curve on travelling paper (in practice, moving galvanometer spot of light traces curve on photographic plate). Rising part of curve shows effect of stimulus; descending part, recovery.
(a) M muscle; A uninjured, B injured ends. E E′ non-polarising electrodes connect A and B with galvanometer G. The stimulus creates a ‘negative variation’ in the resting current. The index linked to the galvanometer needle records a curve on moving paper (in practice, the moving spot of light from the galvanometer traces the curve on a photographic plate). The rising section of the curve indicates the effect of the stimulus; the descending section shows recovery.
(b) O is the zero position of the galvanometer; injury produces a deflection A B; stimulus diminishes this deflection to C; C D is the recovery.
(b) O is the zero position of the galvanometer; injury causes a deflection A B; stimulus reduces this deflection to C; C D represents the recovery.
Two types of response—positive and negative.—It may here be added that though stimulus in general produces a diminution of current of rest, or a negative variation (e.g. muscles and nerves), yet, in certain cases, there is an increase, or positive variation. This is seen in the response of the retina to light. Again, a tissue which normally gives a negative variation may undergo molecular changes, after which it gives a positive variation. Thus Dr. Waller finds that whereas fresh nerve always gives negative variation, stale nerve sometimes gives positive; and that retina, which when fresh gives positive, when stale, exhibits negative variation.
Two types of response—positive and negative.—It’s worth noting that while stimuli generally cause a decrease in the resting current, or a negative variation (for example, in muscles and nerves), there are certain cases where an increase, or positive variation, occurs. This is evident in the retina's response to light. Additionally, a tissue that typically shows a negative variation may undergo molecular changes, after which it displays a positive variation. For instance, Dr. Waller observes that while fresh nerve always shows a negative variation, stale nerve can sometimes show a positive one; and that the retina, which shows a positive variation when fresh, exhibits a negative variation when stale.
I. Negative variation.—Action current from more excited to less excited—cuproid change in the excited—e.g. fresh muscle and nerve, stale retina.
I. Negative variation.—Action moves from a more excited state to a less excited one—cuproid change in the excited—e.g. fresh muscle and nerve, stale retina.
II. Positive variation.—Action current from less excited to more excited—zincoid change in the excited—e.g. stale nerve, fresh retina.[4]
II. Positive variation.—Action runs from less excited to more excited—zincoid change in the excited—e.g. stale nerve, fresh retina.[4]
From this it will be seen that it is the fact of the electrical response of living substances to stimulus that is of essential importance, the sign plus or minus being a minor consideration.
From this, it’s clear that what really matters is how living substances respond to stimuli electrically, while the sign plus or minus is a secondary consideration.
Universal applicability of the electrical mode of response.—This mode of obtaining electrical response is applicable to all living tissues, and in cases like that of muscle, where mechanical response is also available, it is found that the electrical and mechanical records are practically identical.
Universal applicability of the electrical mode of response.—This way of getting an electrical response works for all living tissues, and in situations like that of muscle, where a mechanical response is also possible, it turns out that the electrical and mechanical recordings are basically the same.
The two response-curves seen in the accompanying diagram (fig. 5), and taken from the same muscle by the two methods simultaneously, clearly exhibit this. Thus we see that electrical response can not only take the place of the mechanical record, but has the further advantage of being applicable in cases where the latter cannot be used.
The two response curves shown in the accompanying diagram (fig. 5) and obtained from the same muscle using both methods at the same time clearly demonstrate this. This indicates that electrical response can not only replace the mechanical record but also has the additional advantage of being usable in situations where the mechanical method is not applicable.
Electrical response: A measure of physiological activity.—These electrical changes are regarded as physiological, or characteristic of living tissue, for any conditions which enhance physiological activity also, pari passu, increase their intensity. Again, when the tissue is killed by poison, electrical response disappears, the tissue passing into an irresponsive condition. Anæsthetics, like chloroform, gradually diminish, and finally altogether abolish, electrical response.
Electrical response: A measure of physiological activity.—These electrical changes are considered physiological, or typical of living tissue, because any conditions that boost physiological activity also increase their intensity. Furthermore, when the tissue is killed by poison, the electrical response disappears, and the tissue enters an unresponsive state. Anesthetics, like chloroform, gradually reduce and eventually completely eliminate the electrical response.

Fig. 5.—Simultaneous Record of the Mechanical (M) and (E) Electrical Responses of the Muscle of Frog. (Waller.)
Fig. 5.—Simultaneous Record of the Mechanical (M) and (E) Electrical Responses of the Muscle of Frog. (Waller.)
From these observed facts—that living tissue gives response while a tissue that has been killed does not—it is concluded that the phenomenon of response is peculiar to living organisms.[5] The response phenomena that we have been studying are therefore considered as due to some unknown, super-physical ‘vital’ force and are thus relegated to a region beyond physical inquiry.
From these observed facts—that living tissue responds while dead tissue does not—it is concluded that the phenomenon of response is unique to living organisms.[5] The response phenomena we have been studying are therefore thought to be due to some unknown, non-physical 'vital' force and are thus placed in a realm beyond physical investigation.
Electric response in plants.—But before we proceed to the inquiry as to whether these responses are or are not due to some physical property of matter, and are to be met with even in inorganic substances, it will perhaps be advisable to see whether they are not paralleled by phenomena in the transitional world of plants. We shall thus pass from a study of response in highly complex animal tissues to those given under simpler vital conditions.
Electric response in plants.—Before we look into whether these responses are caused by a physical property of matter, and if they can also be found in inorganic substances, it might be a good idea to examine if they are similar to phenomena in the transitional world of plants. We will move from studying responses in complex animal tissues to those observed under simpler living conditions.
Electric response has been found by Munck, Burdon-Sanderson, and others to occur in sensitive plants. But it would be interesting to know whether these responses were confined to plants which exhibit such remarkable mechanical movements, and whether they could not also be obtained from ordinary plants where visible movements are completely absent. In this connection, Kunkel observed electrical changes in association with the injury or flexion of stems of ordinary plants.[6] My own attempt, however, was directed, not towards the obtaining of a mere qualitative response, but rather to the determination of whether throughout the whole range of response phenomena a parallelism between animal and vegetable could be detected. That is to say, I desired to know, with regard to plants, what was the relation between intensity of stimulus and the corresponding response; what were the effects of superposition of stimuli; whether fatigue was present, and in what manner it influenced response; what were the effects of extremes of temperature on the response; and, lastly, if chemical reagents could exercise any influence in the modification of plant response, as stimulating, anæsthetic, and poisonous drugs have been found to do with nerve and muscle.
Electric responses have been observed by Munck, Burdon-Sanderson, and others in sensitive plants. However, it would be interesting to know if these responses are limited to plants that show such remarkable mechanical movements, and whether they can also be found in ordinary plants where visible movements are completely absent. In this context, Kunkel noted electrical changes related to the injury or bending of stems in regular plants.[6] My own effort, however, was focused not just on obtaining a simple qualitative response, but on determining whether a parallel could be detected between the response phenomena in animals and plants throughout the entire range. In other words, I wanted to understand, regarding plants, the relationship between the intensity of the stimulus and the corresponding response; the effects of combining stimuli; whether fatigue was present and how it influenced the response; the effects of extreme temperatures on the response; and finally, whether chemical reagents could affect the modification of plant responses, just as stimulating, anesthetic, and poisonous drugs do with nerves and muscles.
If it could be proved that the electric response served as a faithful index of the physiological activity of plants, it would then be possible successfully to attack many problems in plant physiology, the solution of which at present offers many experimental difficulties.
If it could be proven that the electrical response accurately reflected the physiological activity of plants, it would be possible to effectively address many issues in plant physiology that currently present significant experimental challenges.
With animal tissues, experiments have to be carried on under many great and unavoidable difficulties. The isolated tissue, for example, is subject to unknown changes inseparable from the rapid approach of death. Plants, however, offer a great advantage in this respect, for they maintain their vitality unimpaired during a very great length of time.
With animal tissues, experiments face many significant and unavoidable challenges. Isolated tissue, for instance, undergoes unknown changes that come with the quick onset of death. However, plants provide a big advantage in this regard, as they can keep their vitality intact for a much longer time.
In animal tissues, again, the vital conditions themselves are highly complex. Those essential factors which modify response can, therefore, be better determined under the simpler conditions which obtain in vegetable life.
In animal tissues, the vital conditions are quite complex. The key factors that influence responses can be more accurately identified under the simpler conditions found in plant life.
In the succeeding chapters it will be shown that the response phenomena are exhibited not only by plants but by inorganic substances as well, and that the responses are modified by various conditions in exactly the same manner as those of animal tissues. In order to show how striking are these similarities, I shall for comparison place side by side the responses of animal tissues and those I have obtained with plants and inorganic substances. For the electric response in animal tissues, I shall take the latest and most complete examples from the records made by Dr. Waller.
In the upcoming chapters, it will be demonstrated that response phenomena are present not just in plants but also in inorganic substances, and that the responses are influenced by various conditions in the same way as those of animal tissues. To highlight these similarities, I will compare the responses of animal tissues with those I have observed in plants and inorganic substances. For the electric response in animal tissues, I will use the most recent and comprehensive examples from the records created by Dr. Waller.
But before we can obtain satisfactory and conclusive results regarding plant response, many experimental difficulties will have to be surmounted. I shall now describe how this has been accomplished.[7]
But before we can get solid and clear results about how plants respond, we need to overcome many experimental challenges. I will now explain how this has been achieved.[7]
FOOTNOTES:
[3] ‘The exciting cause is able to produce a particular molecular rearrangement in the nerve; this constitutes the state of excitation and is accompanied by local electrical changes as an ascertained physical concomitant.’
[3] 'The triggering factor can cause a specific rearrangement of molecules in the nerve; this results in a state of excitation and is associated with local electrical changes that have been confirmed as a physical counterpart.'
‘The excitatory state evoked by stimulus manifests itself in nerve fibres by E.M. changes, and as far as our present knowledge goes by these only. The conception of such an excitable living tissue as nerve implies that of a molecular state which is in stable equilibrium. This equilibrium can be readily upset by an external agency, the stimulus, but the term “stable” expresses the fact that a change in any direction must be succeeded by one of opposite character, this being the return of the living structure to its previous state. Thus the electrical manifestation of the excitatory state is one whose duration depends upon the time during which the external agent is able to upset and retain in a new poise the living equilibrium, and if this is extremely brief, then the recoil of the tissue causes such manifestation to be itself of very short duration.’—Text-book of Physiology, ed. by Schäfer, ii. 453.
‘The excitatory state triggered by a stimulus shows up in nerve fibers through electrical changes, and based on what we currently know, that's the only way. The idea of nerve as an excitable living tissue suggests a molecular state that is in stable balance. This balance can easily be disrupted by an outside force, the stimulus, but the term “stable” indicates that any change in one direction must be followed by a change in the opposite direction, which brings the living structure back to its original state. Therefore, the electrical signs of the excitatory state last as long as the external force can disrupt and keep the living balance in a new position, and if this disruption is very brief, then the tissue’s rebound causes this sign to also be very short-lived.’—Text-book of Physiology, ed. by Schäfer, ii. 453.
[4] I shall here mention briefly one complication that might arise from regarding the current of injury as the current of reference, and designating the response current either positive or negative in relation to it. If this current of injury remained always invariable in direction—that is to say, from the injured to the uninjured—there would be no source of uncertainty. But it is often found, for example in the retina, that the current of injury undergoes a reversal, or is reversed from the beginning. That is to say, the direction is now from the uninjured to the injured, instead of the opposite. Confusion is thus very apt to arise. No such misunderstanding can however occur if we call the current of response towards the more excited positive, and towards the less excited negative.
[4] I want to briefly point out a complication that can come up when we think of the current of injury as the current of reference and label the response current as either positive or negative in relation to it. If this current of injury always stayed the same in direction—that is, from the injured to the uninjured—there wouldn’t be any uncertainty. However, it’s often observed, particularly in the retina, that the current of injury can reverse or be reversed from the start. This means the direction can switch to go from the uninjured to the injured, instead of the other way around. This can easily lead to confusion. However, there won’t be any misunderstanding if we define the current of response going towards the more excited state as positive and towards the less excited state as negative.
[5] ‘The Electrical Sign of Life ... An isolated muscle gives sign of life by contracting when stimulated ... An ordinary nerve, normally connected with its terminal organs, gives sign of life by means of muscle, which by direct or reflex path is set in motion when the nerve trunk is stimulated. But such nerve separated from its natural termini, isolated from the rest of the organism, gives no sign of life when excited, either in the shape of chemical or of thermic changes, and it is only by means of an electrical change that we can ascertain whether or no it is alive ... The most general and most delicate sign of life is then the electrical response.’—Waller, in Brain, pp. 3 and 4. Spring 1900.
[5] ‘The Electrical Sign of Life ... An isolated muscle shows signs of life by contracting when stimulated ... A regular nerve, when connected to its target organs, shows signs of life through muscle, which gets activated, either directly or reflexively, when the nerve is stimulated. However, a nerve that’s separated from its natural endpoints and cut off from the rest of the body shows no signs of life when triggered, either through chemical or thermal changes; it’s only through an electrical change that we can determine if it’s alive ... So, the most common and sensitive indicator of life is the electrical response.’—Waller, in Brain, pp. 3 and 4. Spring 1900.
[6] Kunkel thought the electric disturbance to be due to movement of water through the tissue. It will be shown that this explanation is inadequate.
[6] Kunkel believed the electric disturbance was caused by water moving through the tissue. It will be demonstrated that this explanation is insufficient.
CHAPTER III
ELECTRIC RESPONSE IN PLANTS—METHOD OF NEGATIVE VARIATION
- Negative variation
- —Response recorder
- —Photographic recorder
- —Compensator
- —Means of graduating intensity of stimulus
- —Spring-tapper and torsional vibrator
- —Intensity of stimulus dependent on amplitude of vibration
- —Effectiveness of stimulus dependent on rapidity also.
I shall first proceed to show that an electric response is evoked in plants under stimulation.[8]
I will first demonstrate that plants respond electrically when stimulated.[8]
In experiments for the exhibition of electric response it is preferable to use a non-electrical form of stimulus, for there is then a certainty that the observed response is entirely due to reaction from stimulus, and not, as might be the case with electric stimulus, to mere escape of stimulating current through the tissue. For this reason, the mechanical form of stimulation is the most suitable.
In experiments for the electric response exhibition, it's better to use a non-electrical type of stimulus. This ensures that the response we observe is entirely from the stimulus reaction, rather than, as could happen with electrical stimuli, just the leakage of stimulating current through the tissue. For this reason, mechanical stimulation is the best option.
I find that all parts of the living plant give electric response to a greater or less extent. Some, however, give stronger response than others. In favourable cases, we may have an E.M. variation as high as ·1 volt. It must however be remembered that the response, being a function of physiological activity of the plant, is liable to undergo changes at different seasons of the year. Each plant has its particular season of maximum responsiveness. The leaf-stalk of horse-chestnut, for example, exhibits fairly strong response in spring and summer, but on the approach of autumn it undergoes diminution. I give here a list of specimens which will be found to exhibit fairly good response:
I find that all parts of the living plant show some electric response, though to varying degrees. Some parts, however, respond more strongly than others. In ideal conditions, we can observe an E.M. variation as high as 0.1 volt. It's important to note that the response depends on the physiological activity of the plant, which can change with the seasons. Each plant has its own peak season of responsiveness. For instance, the leaf stalk of the horse-chestnut demonstrates a strong response in spring and summer, but diminishes as autumn approaches. Here’s a list of specimens that show a decent response:
Root.—Carrot (Daucus Carota), radish (Raphanus sativus).
Root.—Carrot (Daucus Carota), radish (Raphanus sativus).
Stem.—Geranium (Pelargonium), vine (Vitis vinifera).
Stem.—Geranium (Pelargonium), grapevine (Vitis vinifera).
Leaf-stalk.—Horse-chestnut (Æsculus Hippocastanum), turnip (Brassica Napus), cauliflower (Brassica oleracea), celery (Apium graveolens), Eucharis lily (Eucharis amazonica).
Leaf-stalk.—Horse-chestnut (Aesculus hippocastanum), turnip (Brassica napus), cauliflower (Brassica oleracea), celery (Apium graveolens), Eucharis lily (Eucharis amazonica).
Flower-stalk.—Arum lily (Richardia africana).
Flower-stalk.—Arum lily (Richardia africana).
Fruit.—Egg-plant (Solanum Melongena).
Vegetable.—Eggplant (Solanum Melongena).
Negative variation.—Taking the leaf-stalk of turnip we kill an area on its surface, say B, by the application of a few drops of strong potash, the area at A being left uninjured. A current is now observed to flow, in the stalk, from the injured B to the uninjured A, as was found to be the case in the animal tissue. The potential difference depends on the condition of the plant, and the season in which it may have been gathered. In the experiment here described (fig. 6, a) its value was ·13 volt.
Negative variation.—By taking the leaf stalk of a turnip, we can damage a specific area on its surface, let's say B, by applying a few drops of strong potash, while leaving the area at A unharmed. A current can now be seen flowing in the stalk from the damaged B to the undamaged A, similar to what was observed in animal tissue. The potential difference depends on the condition of the plant and the season it was harvested. In the experiment described here (fig. 6, a), its value was ·13 volt.

Fig. 6.—(a) Experiment for Exhibiting Electric Response in Plants by Method of Negative Variation. (b) Responses in Leaf-stalk of Turnip to Stimuli of Two Successive Taps, the Second being Stronger.
Fig. 6.—(a) Experiment to Show Electric Response in Plants Using the Method of Negative Variation. (b) Responses in the Leaf-stalk of Turnip to Two Successive Taps, with the Second Being Stronger.
A and B contacts are about 2 cm. apart, B being injured. Plant is stimulated by a tap between A and B. Stimulus acts on both A and B, but owing to injury of B, effect at A is stronger and a negative variation due to differential action occurs.
A and B contacts are about 2 cm apart, with B being injured. The plant reacts when tapped between A and B. The stimulus affects both A and B, but because B is injured, the effect at A is stronger, resulting in a negative variation due to the different responses.
A sharp tap was now given to the stalk, and a sudden diminution, or negative variation, of current occurred, the resting potential difference being decreased by ·026 volt. A second and stronger tap produced a second response, causing a greater diminution of P.D. by ·047 volt (fig. 6, b). The accompanying figure is a photographic record of another set of response-curves (fig. 7). The first three responses are for a given intensity of stimulus, and the next six in response to stimulus nearly twice as strong. It will be noticed that fatigue is exhibited in these responses. Other experiments will be described in the next chapter which show conclusively that the response was not due to any accidental circumstance but was a direct result of stimulation. But I shall first discuss the experimental arrangements and method of obtaining these graphic records.
A sharp tap was given to the stalk, causing a sudden decrease, or negative change, in the current, with the resting potential difference being reduced by 0.026 volts. A second and stronger tap triggered another response, leading to a greater decrease in P.D. by 0.047 volts (fig. 6, b). The accompanying figure is a photo record of another set of response curves (fig. 7). The first three responses correspond to a certain intensity of stimulus, while the next six respond to a stimulus that is nearly twice as strong. You'll notice that these responses show signs of fatigue. Other experiments will be described in the next chapter, which clearly demonstrate that the response wasn’t due to any accidental factors but was a direct result of stimulation. First, though, I will discuss the experimental setup and how these graphic records were obtained.

Fig. 7.—Record of Responses in Plant (Leaf-stalk of Cauliflower) by Method of Negative Variation
Fig. 7.—Record of Responses in Plant (Leaf-stalk of Cauliflower) by Method of Negative Variation
The first three records are for stimulus intensity 1; the next six are for intensity twice as strong; the successive responses exhibit fatigue. The vertical line to the left represents ·1 volt. The record is to be read from right to left.
The first three records are for stimulus intensity 1; the next six are for intensity twice as strong; the following responses show fatigue. The vertical line on the left represents ·1 volt. The record should be read from right to left.
Response recorder.—The galvanometer used is a sensitive dead-beat D’Arsonval. The period of complete swing of the coil under experimental conditions is about 11 seconds. A current of 10-9 ampere produces a deflection of 1 mm. at a distance of 1 metre. For a quick and accurate method of obtaining the records, I devised the following form of response recorder. The curves are obtained directly, by tracing the excursion of the galvanometer spot of light on a revolving drum (fig. 8). The drum, on which is wrapped the paper for receiving the record, is driven by clockwork. Different speeds of revolution can be given to it by adjustment of the clock-governor, or by changing the size of the driving-wheel. The galvanometer spot is thrown down on the drum by the inclined mirror M. The galvanometer deflection takes place at right angles to the motion of the paper. A stylographic pen attached to a carrier rests on the writing surface. The carrier slides over a rod parallel to the drum. As has been said before, the galvanometer deflection takes place parallel to the drum, and as long as the plant rests unstimulated, the pen, remaining coincident with the stationary galvanometer spot on the revolving paper, describes a straight line. If, on stimulation, we trace the resulting excursion of the spot of light, by moving the carrier which holds the pen, the rising portion of the response-curve will be obtained. The galvanometer spot will then return more or less gradually to its original position, and that part of the curve which is traced during the process constitutes the recovery. The ordinate in these curves represents the E.M. variation, and the abscissa the time.
Response recorder.—The galvanometer used is a sensitive dead-beat D’Arsonval. The total swing period of the coil during testing is about 11 seconds. A current of 10-9 ampere produces a deflection of 1 mm at a distance of 1 meter. For a quick and accurate way to get the records, I created the following type of response recorder. The curves are generated directly by tracing the movement of the galvanometer's light spot on a revolving drum (fig. 8). The drum, which has the paper for recording wrapped around it, is powered by clockwork. Different speeds of rotation can be set by adjusting the clock governor or by changing the size of the driving wheel. The galvanometer spot is projected onto the drum by the angled mirror M. The galvanometer deflection occurs at right angles to the movement of the paper. A stylus pen attached to a carrier sits on the writing surface. The carrier slides over a rod that runs parallel to the drum. As mentioned earlier, the galvanometer deflection happens parallel to the drum, and as long as the plant remains unstimulated, the pen stays aligned with the stationary galvanometer spot on the rotating paper, tracing a straight line. If we trace the resulting movement of the light spot upon stimulation by moving the carrier with the pen, we will obtain the rising part of the response curve. The galvanometer spot will then gradually return to its original position, and the part of the curve traced during this return constitutes the recovery. The vertical axis in these curves represents the E.M. variation, while the horizontal axis represents time.

We can calibrate the value of the deflection by applying a known E.M.F. to the circuit from a compensator, and noting the deflection which results. The speed of the clock is previously adjusted so that the recording surface moves exactly through, say, one inch a minute. Of course this speed can be increased to suit the particular experiment, and in some it is as high as six inches a minute. In this simple manner very accurate records may be made. It has the additional advantage that one is able at once to see whether the specimen is suitable for the purpose of investigation. A large number of records might be taken by this means in a comparatively short time.
We can adjust the deflection value by applying a known E.M.F. to the circuit from a compensator and observing the resulting deflection. The clock speed is set in advance so that the recording surface moves exactly one inch per minute. Of course, this speed can be increased depending on the specific experiment, and in some cases, it goes up to six inches per minute. This straightforward method allows for very accurate records. It also has the added benefit of letting you quickly determine whether the specimen is suitable for investigation. With this approach, you can collect a large number of records in a relatively short time.
Photographic recorder.—Or the records may be made photographically. A clockwork arrangement moves a photographic plate at a known uniform rate, and a curve is traced on the plate by the moving spot of light. All the records that will be given are accurate reproductions of those obtained by one of these two methods. Photographic records are reproduced in white against a black background.
Photographic recorder.—Records can also be created using photography. A mechanical setup moves a photographic plate at a consistent speed, and a light spot traces a curve on the plate. All the records provided here are precise reproductions of those made using either of these two methods. Photographic records appear in white on a black background.
Compensator.—As the responses are on variation of current of injury, and as the current of injury may be strong, and throw the spot of light beyond the recording surface, a potentiometer balancing arrangement may be used (fig. 9), by which the P.D. due to injury is exactly compensated; E.M. variations produced by stimulus are then taken in the usual manner. This compensating arrangement is also helpful, as has been said before, for calibrating the E.M. value of the deflection.
Compensator.—Since the responses depend on the variation of the injury current, and since the injury current can be strong enough to move the spot of light beyond the recording area, a potentiometer balancing setup may be used (fig. 9). This setup precisely compensates for the potential difference caused by the injury; the E.M. variations produced by the stimulus are then measured in the usual way. This compensating setup is also useful, as previously mentioned, for calibrating the E.M. value of the deflection.

Fig. 9.—The Compensator
Fig. 9.—The Compensator
A B is a stretched wire with added resistances R and R′. S is a storage cell. When the key K is turned to the right one scale division = ·001 volt, when turned to the left one scale division = ·01 volt. P is the plant.
A B is a stretched wire with added resistances R and R′. S is a storage cell. When the switch K is turned to the right, one scale division equals 0.001 volt; when turned to the left, one scale division equals 0.01 volt. P is the plant.
Means of graduating the intensity of stimulus.—One of the necessities in connection with quantitative measurements is to be certain that the intensity of successive stimuli is (1) constant, or (2) capable of gradual increase by known amounts. No two taps given by the hand can be made exactly alike. I have therefore devised the two following methods of stimulation, which have been found to act satisfactorily.
Ways to Gradually Increase the Intensity of a Stimulus.—One of the key requirements for quantitative measurements is to ensure that the intensity of consecutive stimuli is (1) consistent, or (2) able to gradually increase by specific amounts. No two taps from the hand can be exactly the same. Therefore, I have developed the following two methods of stimulation, which have proven to be effective.
The spring-tapper.—This consists (fig. 10) of the spring proper (S), the attached rod (R) carrying at its end the tapping-head (T). A projecting rod—the lifter (L)—passes through S R. It is provided with a screw-thread, by means of which its length, projecting downwards, is regulated. This fact, as we shall see, is made to determine the height of the stroke. (C) is a cogwheel. As one of the spokes of the cogwheel is rotated past (L), the spring is lifted and released, and (T) delivers a sharp tap. The height of the lift, and therefore the intensity of the stroke, is measured by means of a graduated scale. We can increase the intensity of the stroke through a wide range (1) by increasing the projecting length of the lifter, and (2) by shortening the length of spring by a sliding catch. We may give isolated single taps or superpose a series in rapid succession according as the wheel is rotated slow or fast. The only disadvantage of the tapping method of stimulation is that in long-continued experiment the point struck is liable to be injured. The vibrational mode of stimulation to be presently described labours under no such disadvantage.
The spring-tapper.—This consists (fig. 10) of the spring itself (S), the attached rod (R) that carries the tapping head (T) at its end. A protruding rod—the lifter (L)—goes through S R. It has a screw thread that allows us to adjust its length extending downward. This feature will help determine the height of the stroke. (C) is a cogwheel. As one of the spokes of the cogwheel spins past (L), the spring is lifted and released, causing (T) to deliver a sharp tap. The height of the lift, and thus the intensity of the stroke, is measured using a graduated scale. We can increase the intensity of the stroke over a wide range (1) by extending the length of the lifter, and (2) by shortening the spring's length with a sliding catch. We can produce single taps or a series of quick taps depending on whether the wheel is turned slowly or quickly. The only drawback of the tapping method of stimulation is that prolonged use may damage the area being tapped. The vibrational method of stimulation that will be described next does not have this issue.

Fig. 11.—The Torsional Vibrator
Fig. 11.—The Torsional Vibrator
Plant P is securely held by a vice V. The two ends are clamped by holders C C′. By means of handles H H′, torsional vibration may be imparted to either the end A or end B of the plant. The end view (b) shows how the amplitude of vibration is predetermined by means of movable stops S S′.
Plant P is firmly fixed in a vice V. The two ends are gripped by holders C C′. Using handles H H′, torsional vibration can be applied to either end A or end B of the plant. The end view (b) illustrates how the vibration amplitude is set using adjustable stops S S′.
Vibrational stimulus.—I find that torsional vibration affords another very effective method of stimulation (fig. 11). The plant-stalk may be fixed in a vice (V), the free ends being held in tubes (C C′), provided with three clamping jaws. A rapid torsional vibration[9] may now be imparted to the stalk by means of the handle (H). The amplitude of vibration, which determines the intensity of stimulus, can be accurately measured by the graduated circle. The amplitude of vibration may be predetermined by means of the sliding stops (S S′).
Vibrational stimulus.—I’ve found that torsional vibration offers another very effective method of stimulation (fig. 11). The plant stalk can be secured in a vice (V), with the free ends held in tubes (C C′) that are equipped with three clamping jaws. A quick torsional vibration[9] can now be applied to the stalk using the handle (H). The vibration's amplitude, which determines the intensity of the stimulus, can be accurately measured using the graduated circle. The vibration amplitude can be set in advance using the sliding stops (S S′).
Intensity of stimulus dependent on amplitude of vibration.—I shall now describe an experiment which shows that torsional vibration is as effective as stimulation by taps, and that its stimulating intensity increases, length of stalk being constant, with amplitude of vibration. It is of course obvious that if the length of the specimen be doubled, the vibration, in order to produce the same effect, must be through twice the angle. I took a leaf-stalk of turnip and fixed it in the torsional vibrator. I then took record of responses to two successive taps, the intensity of one being nearly double that of the other. Having done this, I applied to the same stalk two successive torsional vibrations of 45° and 67° respectively. These successive responses to taps and torsional vibrations are given in fig. 12, and from them it will be seen that these two modes of stimulation may be used indifferently, with equal effect. The vibrational method has the advantage over tapping, that, while with the latter the stimulus is somewhat localised, with vibration the tissue subjected to stimulus is uniformly stimulated throughout its length.
Intensity of stimulus dependent on amplitude of vibration.—I will now describe an experiment that demonstrates that torsional vibration is just as effective as stimulation from taps, and that its stimulating intensity increases, with the length of the stalk remaining constant, as the amplitude of vibration increases. It's clear that if the length of the specimen is doubled, the vibration must move through twice the angle to produce the same effect. I took a turnip leaf stalk and fixed it in the torsional vibrator. I then recorded responses to two successive taps, one being nearly double the intensity of the other. After this, I applied two successive torsional vibrations of 45° and 67° respectively to the same stalk. The responses to the taps and torsional vibrations are provided in fig. 12, and from them, you can see that these two methods of stimulation can be used interchangeably with equal effectiveness. The vibrational method has the advantage over tapping in that, while tapping produces a somewhat localized stimulus, vibration stimulates the tissue evenly along its entire length.
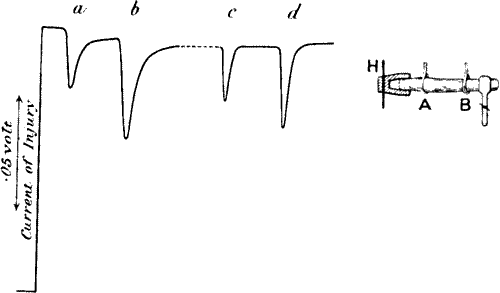
Fig. 12.—Response in Plant to Mechanical Tap or Vibration
Fig. 12.—Response in Plants to Mechanical Tap or Vibration
The end B is injured. A tap was given between A and B and this gave the response-curve a. A stronger tap gave the response b. By means of the handle H, a torsional vibration of 45° was now imparted, this gave the response c. Vibration through 67° gave d.
The end B is injured. A tap was made between A and B, which produced the response curve a. A stronger tap resulted in the response b. Using the handle H, a torsional vibration of 45° was applied, resulting in the response c. A vibration of 67° produced d.
Effectiveness of stimulus dependent on rapidity also. In order that successive stimuli may be equally effective another point has to be borne in mind. In all cases of stimulation of living tissue it is found that the effectiveness of a stimulus to arouse response depends on the rapidity of the onset of the disturbance. It is thus found that the stimulus of the ‘break’ induction shock, on a muscle for example, is more effective, by reason of its greater rapidity, than the ‘make’ shock. So also with the torsional vibrations of plants, I find response depending on the quickness with which the vibration is effected. I give below records of successive stimuli, given by vibrations through the same amplitude, but delivered with increasing rapidity (fig. 13).
Effectiveness of stimulus also depends on speed. For successive stimuli to be equally effective , we need to keep another point in mind. In all instances of stimulating living tissue, it's observed that how effective a stimulus is at eliciting a response relies on how quickly the disturbance occurs. For example, the stimulus from a ‘break’ induction shock to a muscle is more effective due to its faster onset than a ‘make’ shock. Similarly, in the torsional vibrations of plants, I notice that the response is influenced by how quickly the vibration happens. Below, I present records of successive stimuli that were administered with the same amplitude but at increasing speeds (fig. 13).

Fig. 13.—Influence of Suddenness on the Efficiency of Stimulus
Fig. 13.—Effect of Suddenness on the Effectiveness of Stimulus
The curves a, b, c, d, are responses to vibrations of the same amplitude, 30°. In a the vibration was very slow; in b it was less slow; it was rapid in c, and very rapid in d.
The curves a, b, c, d show responses to vibrations of the same amplitude, 30°. In a, the vibration was very slow; in b, it was slightly less slow; it was fast in c, and very fast in d.
Thus if we wish to maintain the effective intensity of stimulus constant we must meet two conditions: (1) The amplitude of vibration must be kept the same. This is done by means of the graduated circle. (2) The vibration period must be kept the same. With a little practice, this requirement is easily fulfilled.
Thus, if we want to keep the effective intensity of the stimulus constant, we need to meet two conditions: (1) The amplitude of vibration must remain the same. This is achieved using the graduated circle. (2) The vibration period must also be consistent. With a bit of practice, this requirement is easily met.
The uniformity of stimulation which is thus attained solves the great difficulty of obtaining reliable quantitative values, by whose means alone can rigorous demonstration of the phenomena we are studying become possible.
The consistency of stimulation achieved this way addresses the significant challenge of getting reliable quantitative values, which are essential for rigorously demonstrating the phenomena we are examining.
FOOTNOTES:
[8] A preliminary account of Electric Response in Plants was given at the end of my paper on ‘Electric Response of Inorganic Substances’ read before the Royal Society on June 6, 1901; also at the Friday Evening Discourse, Royal Institution, May 10, 1901. A more complete account is given in my paper on ‘Electric Response in Ordinary Plants under Mechanical Stimulus’ read before the Linnean Society March 20, 1902.
[8] I provided an initial overview of Electric Response in Plants at the end of my paper on ‘Electric Response of Inorganic Substances’ presented to the Royal Society on June 6, 1901; and also during the Friday Evening Discourse at the Royal Institution on May 10, 1901. A more detailed account is presented in my paper on ‘Electric Response in Ordinary Plants under Mechanical Stimulus,’ which was delivered to the Linnean Society on March 20, 1902.
I thank the Royal Society and the Linnean Society for permission to reproduce some of my diagrams published in their Proceedings.—J. C. B.
I thank the Royal Society and the Linnean Society for allowing me to reproduce some of my diagrams published in their Proceedings.—J. C. B.
[9] By this is meant a rapid to-and-fro or complete vibration. In order that successive responses should be uniform it is essential that there should be no resultant twist, i.e. the plant at the end of vibration should be in exactly the same condition as at the beginning.
[9] This refers to a quick back-and-forth or full vibration. To ensure that each response is consistent, it's crucial that there's no twisting effect. This means that by the end of the vibration, the plant should be in the exact same condition as it was at the start.
CHAPTER IV
ELECTRIC RESPONSE IN PLANTS—BLOCK METHOD
- Method of block
- —Advantages of block method
- —Plant response a physiological phenomenon
- —Abolition of response by anæsthetics and poisons
- —Abolition of response when plant is killed by hot water.
I shall now proceed to describe another and independent method which I devised for obtaining plant response. It has the advantage of offering us a complementary means of verifying the results found by the method of negative variation. As it is also, in itself, for reasons which will be shown later, a more perfect mode of inquiry, it enables us to investigate problems which would otherwise have been difficult to attempt.
I will now describe another separate method I created to observe plant responses. This method adds an extra way to check the results we found using the method of negative variation. Additionally, for reasons I will explain later, it is a more effective way of inquiry, allowing us to explore problems that would have been hard to tackle otherwise.
When electrolytic contacts are made on the uninjured surfaces of the stalk at A and B, the two points, being practically similar in every way, are iso-electric, and little or no current will flow in the galvanometer. If now the whole stalk be uniformly stimulated, and if both ends A and B be equally excited at the same moment, it is clear that there will still be no responsive current, owing to balancing action at the two ends. This difficulty as regards the obtaining of response was overcome in the method of negative variation, where the excitability of one end was depressed by chemical reagents or injury, or abolished by excessive tempera ture. On stimulating the stalk there was produced a greater excitation at A than at B, and a current of action was then observed to flow in the stalk from the more excited A to the less excited B (fig. 6).
When electrolytic contacts are made on the uninjured surfaces of the stalk at A and B, the two points, being nearly identical in every way, are iso-electric, and little to no current will flow in the galvanometer. If the entire stalk is stimulated uniformly and both ends A and B are evenly excited at the same time, it’s evident that there will still be no current flow because the action at both ends balances out. This challenge in getting a response was resolved in the method of negative variation, where the excitability of one end was reduced by chemical substances or injury, or eliminated by very high temperatures. When the stalk was stimulated, a stronger excitation at A compared to B resulted in a current of action flowing in the stalk from the more excited A to the less excited B (fig. 6).
But we can cause this differential action to become evident by another means. For example, if we produce a block, by clamping at C between A and B (fig. 14, a), so that the disturbance made at A by tapping or vibration is prevented from reaching B, we shall then have A thrown into a relatively greater excitatory condition than B. It will now be found that a current of action flows in the stalk from A to B, that is to say, from the excited to the less excited. When the B end is stimulated, there will be a reverse current (fig. 14, b).
But we can make this differential action clear in another way. For example, if we create a block by clamping at C between A and B (fig. 14, a), preventing the disturbance at A from reaching B through tapping or vibration, we will find that A is in a relatively more excited state than B. It will then be observed that an action current flows in the stalk from A to B, meaning from the more excited to the less excited. When the B end is stimulated, a reverse current will occur (fig. 14, b).

Fig. 14.—The Method of Block
Fig. 14.—Block Method
(a) The plant is clamped at C, between A and B.
(a) The plant is secured at C, between A and B.
(b) Responses obtained by alternately stimulating the two ends. Stimulation of A produces upward response; of B gives downward response.
(b) Responses obtained by alternating stimulation at the two ends. Stimulating A causes an upward response; stimulating B causes a downward response.
We have in this method a great advantage over that of negative variation, for we can always verify any set of results by making corroborative reversal experiments.
We have a big advantage with this method compared to negative variation, because we can always confirm any set of results by conducting supporting reversal experiments.
By the method of injury again, one end is made initially abnormal, i.e. different from the condition which it maintains when intact. Further, inevitable changes will proceed unequally at the injured and uninjured ends, and the conditions of the experiment may thus undergo unknown variations. But by the block method which has just been described, there is no injury, the plant is normal throughout, and any physiological change (which in plants will be exceedingly small during the time of the experiment) will affect it as a whole.
By using the injury method again, one end is made deliberately abnormal, meaning it’s different from the state it remains in when undamaged. Additionally, unavoidable changes will occur unevenly at the injured and uninjured ends, leading to unknown variations in the conditions of the experiment. However, with the block method that has just been explained, there is no injury; the plant remains normal throughout, and any physiological change (which will be very slight in plants during the experiment) will affect the entire plant.

Fig. 15.—Response in Plant (from the Stimulated A to Unstimulated B) Completely Immersed Under Water
Fig. 15.—Response in Plant (from the Stimulated A to Unstimulated B) Fully Submerged in Water
The leaf-stalk is clamped securely in the middle with the cork C, inside the tube T, which is filled with water, the plant being completely immersed. Moistened threads in connection with the two non-polarisable electrodes are led to the side tubes t t′. One end of the stalk is held in ebonite forceps and vibrated. A current of response is found to flow in the stalk from the excited A to the unexcited B, and outside, through the liquid, from B to A. A portion of this current, flowing through the side tubes t t′, produces deflection in the galvanometer.
The leaf stalk is securely clamped in the middle with the cork C, inside the tube T, which is filled with water, so the plant is completely submerged. Moistened threads connected to the two non-polarizable electrodes are led to the side tubes t t′. One end of the stalk is held in ebonite forceps and vibrated. A response current is found to flow in the stalk from the excited A to the unexcited B, and outside, through the liquid, from B to A. A portion of this current, flowing through the side tubes t t′, causes deflection in the galvanometer.
Plant response a physiological or vital response.—I now proceed to a demonstration of the fact that whatever be the mechanism by which they are brought about, these plant responses are physiological in their character. As the investigations described in the next few chapters will show, they furnish an accurate index of physiological activity. For it will be found that, other things being equal, whatever tends to exalt or depress the vitality of the plant tends also to increase or diminish its electric response. These E.M. effects are well marked, and attain considerable value, rising sometimes, as has been said before, to as much as ·1 volt or more. They are proportional to the intensity of stimulus.
Plant responses are physiological or vital reactions.—I will now demonstrate that, regardless of the mechanisms involved, these plant responses are fundamentally physiological. The investigations outlined in the following chapters will show that they provide an accurate measure of physiological activity. It will be found that, all else being equal, anything that boosts or reduces the vitality of the plant also tends to increase or decrease its electrical response. These electrical effects are significant and can reach values of up to 1 volt or more, as mentioned before. They are proportional to the intensity of the stimulus.
It need hardly be added that special precautions are taken to avoid shifting of contacts. Variation of contact, however, could not in any case account for repeated transient responses to repeated stimuli, when contact is made on iso-electric surfaces. Nor could it in any way explain the reversible nature of these responses, when A and B are stimulated alternately. These responses are obtained in the plants even when completely immersed in water, as in the experimental arrangement (fig. 15). It will be seen that in this case, where there could be no possibility of shifting of contact, or variation of surface, there is still the usual current of response.
It barely needs mentioning that special precautions are taken to avoid contact shifting. However, variations in contact couldn't possibly explain the repeated short-term responses to repeated stimuli when contact is made on iso-electric surfaces. Nor could it explain the reversible nature of these responses when A and B are stimulated alternately. These responses occur in the plants even when fully submerged in water, as shown in the experimental setup (fig. 15). It's clear that in this situation, where there's no chance of contact shifting or surface variation, there is still the usual response current.
I shall describe here a few crucial experiments only, in proof of the physiological character of electric response. The test applied by physiologists, in order to discriminate as to the physiological nature of response, consists in finding out whether the response is diminished or abolished by the action of anæsthetics, poisons, and excessively high temperature, which are known to depress or destroy vitality.
I will describe just a few key experiments here to prove the physiological nature of electric response. The test used by physiologists to determine the physiological nature of the response involves checking if the response is reduced or eliminated by the effects of anesthetics, poisons, and extremely high temperatures, which are known to lower or eliminate vitality.
I shall therefore apply these same tests to plant responses.
I will apply these same tests to how plants respond.
Effect of anæsthetics and poisons.—Ordinary anæsthetics, like chloroform, and poisons, like mercuric chloride, are known to produce a profound depression or abolish all signs of response in the living tissue. For the purpose of experiment, I took two groups of stalks, with leaves attached, exactly similar to each other in every respect. In order that the leaf-stalks might absorb chloroform I dipped their cut ends in chloroform-water, a certain amount of which they absorbed, the process being helped by the transpiration from the leaves. The second group of stalks was placed simply in water, in order to serve for control experiment. The narcotic action of chloroform, finally culminating in death, soon became visually evident. The leaves began to droop, a peculiar death-discolouration began to spread from the mid rib along the venation of the leaves. Another peculiarity was also observed. The aphides feeding on the leaves died even before the appearance of the discoloured patches, whereas on the leaves of the stalks placed in water these little creatures maintained their accustomed activity, nor did any discolouration occur. In order to study the effect of poison, another set was placed in water containing a small quantity of mercuric chloride. The leaves here underwent the same change of appearance, and the aphides met with the same untimely fate, as in the case of those subjected to the action of chloroform. There was hardly any visible change in the appearance of the stalks themselves, which were to all outer seeming as living as ever, indications of death being apparent only on the leaf surfaces. I give below the results of several sets of experiments, from which it would appear that whereas there was strong normal response in the group of stalks kept in water, there was practically a total abolition of all response in those anæsthetised or poisoned.
Effect of anesthetics and poisons.—Common anesthetics, like chloroform, and poisons, such as mercuric chloride, are known to cause a significant depression or eliminate all signs of response in living tissue. For the experiment, I took two groups of stalks, with leaves attached, that were identical in every way. To allow the leaf-stalks to absorb chloroform, I dipped their cut ends in chloroform-water, which they absorbed, aided by the transpiration from the leaves. The second group of stalks was placed simply in water to serve as a control experiment. The narcotic effect of chloroform, which ultimately led to death, became visually clear shortly after. The leaves began to droop, and a distinct death discoloration started spreading from the midrib along the veins of the leaves. Another notable observation was that the aphids feeding on the leaves died even before the discolored patches appeared, while on the leaves of the stalks placed in water, these small creatures continued their usual activity, and no discoloration occurred. To study the effect of poison, another set was placed in water that contained a small amount of mercuric chloride. The leaves showed the same change in appearance, and the aphids met the same early demise as those subjected to chloroform. There was hardly any visible change in the appearance of the stalks themselves, which looked just as alive as ever, with signs of death evident only on the leaf surfaces. Below are the results of several sets of experiments, showing that while there was strong normal response in the group of stalks kept in water, there was practically a total loss of all response in those anesthetized or poisoned.
Experiments on the effect of anæsthetics and poisons. A batch of ten leaf-stalks of plane-tree was placed with the cut ends in water, and leaves in air; an equal number was immersed in chloroform-water; a third batch was placed in 5 per cent. solution of mercuric chloride.
Experiments on the effect of anesthetics and poisons. A group of ten plane tree leaf stalks was put in water with the cut ends submerged, while the leaves were exposed to air; an equal number was soaked in chloroform-water; a third group was placed in a 5 percent solution of mercuric chloride.
Similarly a batch of three horse-chestnut leaf-stalks was put in water, another batch in chloroform-water, and a third batch in mercuric chloride solution.
Similarly, a group of three horse-chestnut leaf stalks was placed in water, another group in chloroform-water, and a third group in mercuric chloride solution.
These results conclusively prove the physiological nature of the response.
These results clearly demonstrate the physiological nature of the response.
I shall in a succeeding chapter give a continuous series of response-curves showing how, owing to progressive death from the action of poison, the responses undergo steady diminution till they are completely abolished.
I will provide a continuous series of response curves in a later chapter, demonstrating how, due to gradual death from poisoning, the responses decrease steadily until they are entirely eliminated.
Effect of high temperature.—It is well known that plants are killed when subjected to high temperatures. I took a stalk, and, using the block method, with torsional vibration as the stimulus, obtained strong responses at both ends A and B. I then immersed the same stalk for a short time in hot water at about 65° C., and again stimulated it as before. But at neither A nor B could any response now be evoked. As all the external conditions were the same in the first and second parts of this experiment, the only difference being that in one the stalk was alive and in the other killed, we have here further and conclusive proof of the physiological character of electric response in plants.
Effect of High Temperature.—It's well known that plants die when exposed to high temperatures. I took a stalk and used the block method with torsional vibration as the stimulus, which produced strong responses at both ends A and B. I then dipped the same stalk in hot water at about 65° C. for a short time and stimulated it again as before. However, at neither A nor B could I elicit any response this time. Since all external conditions were the same in both parts of this experiment, with the only difference being that in one case the stalk was alive and in the other it was dead, we have further and definitive proof of the physiological nature of electric response in plants.
The same facts may be demonstrated in a still more striking manner by first obtaining two similar but opposite responses in a fresh stalk, at A and B, and then killing one half, say B, by immersing only that half of the stalk in hot water. The stalk is replaced in the apparatus, and it is now found that whereas the A half gives strong response, the end B gives none.
The same facts can be shown in an even more compelling way by first getting two similar but opposite reactions in a fresh stalk, at A and B, and then killing one half, for example, B, by immersing just that part of the stalk in hot water. The stalk is put back in the apparatus, and now it's found that while the A half produces a strong response, the end B shows no reaction.
In the experiments on negative variation, it was tacitly assumed that the variation is due to a differential action, stimulus producing a greater excitation at the uninjured than at the injured end. The block method enables us to test the correctness of this assumption. The B end of the stalk is injured or killed by a few drops of strong potash, the other end being uninjured. There is a clamp between A and B. The end A is stimulated and a strong response is obtained. The end B is now stimulated, and there is little or no response. The block is now removed and the plant stimulated throughout its length. Though the stimulus now acts on both ends, yet, owing to the irresponsive condition of B, there is a resultant response, which from its direction is found to be due to the responsive action of A. This would not have been the case if the end B had been uninjured. We have thus experimentally verified the assumption that in the same tissue an uninjured portion will be thrown into a greater excitatory state than an injured, by the action of the same stimulus.
In the experiments involving negative variation, it was assumed that the variation is due to a differential action, where the stimulus causes more excitation at the uninjured end compared to the injured end. The block method allows us to test this assumption. The B end of the stalk is damaged or killed with a few drops of strong potash, while the other end remains uninjured. A clamp is placed between A and B. When the end A is stimulated, it produces a strong response. However, when the end B is stimulated, there is little to no response. The block is then removed, and the plant is stimulated along its entire length. Although the stimulus is now applied to both ends, the unresponsive state of B results in a response that, based on its direction, is determined to be due to the responsive action of A. This wouldn’t have happened if the end B had been uninjured. Thus, we have experimentally confirmed the assumption that within the same tissue, an uninjured section will be more easily excited by the same stimulus than an injured one.
CHAPTER V
PLANT RESPONSE—ON THE EFFECTS OF SINGLE STIMULUS AND OF SUPERPOSED
STIMULI
- Effect of single stimulus
- —Superposition of stimuli
- —Additive effect
- —Staircase effect
- —Fatigue
- —No fatigue when sufficient interval between stimuli
- —Apparent fatigue when stimulation frequency is increased
- —Fatigue under continuous stimulation.
Effect of single stimulus.—In a muscle a single stimulus gives rise to a single twitch which may be recorded either mechanically or electrically. If there is no fatigue, the successive responses to uniform stimuli are exactly similar. Muscle when strongly stimulated often exhibits fatigue, and successive responses therefore become feebler and feebler. In nerves, however, there is practically no fatigue and successive records are alike. Similarly, in plants, we shall find some exhibiting marked fatigue and others very little.
Effect of single stimulus.—In a muscle, a single stimulus causes a single twitch that can be recorded either mechanically or electrically. If there is no fatigue, the responses to the same stimuli are identical. When a muscle is strongly stimulated, it often shows fatigue, leading to weaker and weaker responses. In nerves, however, there’s almost no fatigue, and the responses are consistent. Similarly, in plants, some show significant fatigue while others show very little.


Fig. 17.—Fusion of Effect of Rapidly Succeeding Stimuli
Fig. 17.—Combination of Effects from Quickly Following Stimuli
(a) in muscle; (b) in carrot.
(a) in muscle; (b) in carrot.
Superposition of stimuli.—If instead of a single stimulus a succession of stimuli be superposed, it happens that a second shock is received before recovery from the first has taken place. Individual effects will then become more or less fused. When the frequency is sufficiently increased, the intermittent effects are fused, and we find an almost unbroken curve. When for example the muscle attains its maximum contraction (corresponding to the frequency and strength of stimuli) it is thrown into a state of complete tetanus, in which it appears to be held rigid. If the rapidity be not sufficient for this, we have the jagged curve of incomplete tetanus. If there is not much fatigue, the upper part of the tetanic curve is approximately horizontal, but in cases where fatigue sets in quickly, the fact is shown by the rapid decline of the curve. With regard to all these points we find strict parallels in plant response. In cases where there is no fatigue, the successive responses are identical (fig. 16). With superposition of stimuli we have fusion of effects, analogous to the tetanus of muscle (fig. 17). And lastly, the influence of fatigue in plants is to produce a modification of response-curve exactly similar to that of muscle (see below). One effect of superposition of stimuli may be mentioned here.
Superposition of stimuli.—If instead of just one stimulus multiple stimuli are combined, a second shock can be received before fully recovering from the first. The individual effects will then merge to some extent. When the frequency is high enough, the intermittent effects blend together, creating an almost continuous curve. For instance, when a muscle reaches its maximum contraction (based on the frequency and strength of stimuli), it enters a state of complete tetanus, appearing rigid. If the speed isn't fast enough for this, we see the jagged curve of incomplete tetanus. If there's not much fatigue, the top of the tetanic curve remains fairly flat, but if fatigue sets in quickly, this is indicated by a rapid drop in the curve. In all these aspects, we see clear parallels in how plants respond. When there's no fatigue, the successive responses are identical (fig. 16). With the superposition of stimuli, effects fuse together, similar to muscle tetanus (fig. 17). Finally, the impact of fatigue in plants results in a change in the response curve that is exactly like that of muscle (see below). One effect of combining stimuli can be mentioned here.

Fig. 18.—Additive Effect
Additive Effect
(a) A single stimulus of 3° vibration produced little or no effect, but the same stimulus when rapidly superposed thirty times, produced the large effect (b). (Leaf-stalk of turnip.)
(a) A single vibration at 3° created little to no effect, but the same vibration when applied rapidly thirty times created a significant effect (b). (Leaf-stalk of turnip.)
Additive effect.—It is found in animal responses that there is a minimum intensity of stimulus, below which no response can be evoked. But even a sub-minimal stimulus will, though singly ineffective, become effective by the summation of several. In plants, too, we obtain a similar effect, i.e. the summation of single ineffective stimuli produces effective response (fig. 18).
Additive effect.—Research shows that in animals, there’s a minimum level of stimulus required to trigger a response. However, even a stimulus that’s below this minimum can still lead to a response when combined with several others. We see a similar phenomenon in plants, where the combination of several individual weak stimuli can create a strong response (fig. 18).
Staircase effect.—Animal tissues sometimes exhibit what is known as the ‘staircase effect,’ that is to say, the heights of successive responses are gradually increased, though the stimuli are maintained constant. This is exhibited typically by cardiac muscle, though it is not unknown even in nerve. The cause is obscure, but it seems to depend on the condition of the tissue. It appears as if the molecular sluggishness of tissue were in these cases only gradually removed under stimulation, and the increased effects were due to increased molecular mobility. Whatever be the explanation, I have sometimes observed the same staircase effect in plants (fig. 19).
Staircase effect.—Animal tissues sometimes show what’s called the ‘staircase effect,’ which means that the heights of successive responses gradually increase even though the stimuli remain constant. This is typically seen in cardiac muscle, but it's also observed in nerve tissue. The reason for this is unclear, but it seems related to the condition of the tissue. It appears that the molecular sluggishness in the tissue is only gradually lifted with stimulation, and the increased effects are a result of heightened molecular mobility. Whatever the reason, I have occasionally noticed the same staircase effect in plants (fig. 19).

Fatigue.—It is assumed that in living substances like muscle, fatigue is caused by the break down or dissimilation of tissue by stimulus. And till this waste is repaired by the process of building-up or assimilation, the functional activity of the tissue will remain below par. There may also be an accumulation of the products of dissimilation—‘the fatigue stuffs’—and these latter may act as poisons or chemical depressants.
Fatigue.—It’s believed that in living tissues like muscle, fatigue is caused by the breakdown or dissimilation of tissue due to stimulation. Until this damage is repaired through the process of rebuilding or assimilation, the tissue's functional activity will stay below its normal level. Additionally, there might be a buildup of dissimilation byproducts—often referred to as ‘fatigue stuff’—and these substances could act as toxins or chemical depressants.
In an animal it is supposed that the nutritive blood supply performs the two-fold task of bringing material for assimilation and removing the fatigue products, thus causing the disappearance of fatigue. This explanation, however, is shown to be insufficient by the fact that an excised bloodless muscle recovers from fatigue after a short period of rest. It is obvious that here the fatigue has been removed by means other than that of renewed assimilation and removal of fatigue products by the circulating blood. It may therefore be instructive to study certain phases of fatigue exhibited under simpler conditions in vegetable tissue, where the constructive processes are in abeyance, and there is no active circulation for the removal of fatigue products.
In animals, it's believed that the nutrient-rich blood supply serves two main purposes: delivering materials for processing and eliminating waste products related to fatigue, thereby alleviating tiredness. However, this explanation falls short considering that a severed, bloodless muscle can recover from fatigue after a short rest. It's clear that in this case, fatigue is eliminated through mechanisms other than fresh nutrient delivery and waste removal by circulating blood. Therefore, it might be helpful to examine certain aspects of fatigue observed under simpler conditions in plant tissues, where growth processes are on hold, and there isn't an active circulation to eliminate fatigue byproducts.
It has been said before that the E.M. variation caused by stimulus is the concomitant of a disturbance of the molecules of the responsive tissues from their normal equilibrium, and that the curve of recovery exhibits the restoration of the tissue to equilibrium.
It has been said before that the E.M. variation caused by a stimulus is linked to a disruption of the molecules in the responsive tissues from their normal balance, and that the recovery curve shows the tissue returning to that balance.
No fatigue when sufficient interval between successive stimuli.—We may thus gather from a study of the response-curve some indication of the molecular distortion experienced by the excited tissue. Let us first take the case of an experiment whose record is given in fig. 20, a. It will be seen from that curve that one minute after the application of stimulus there is a complete recovery of the tissue; the molecular condition is exactly the same at the end of recovery as in the beginning of stimulation. The second and succeeding response-curves therefore are exactly similar to the first, provided a sufficient interval has been allowed in each case for complete recovery. There is, in such a case, no diminution in intensity of response, that is to say, no fatigue.
No fatigue when there’s enough time between stimuli.—From examining the response curve, we can get an idea of the molecular changes happening in the stimulated tissue. First, let's look at an experiment recorded in fig. 20, a. The curve shows that one minute after the stimulus, the tissue fully recovers; the molecular state is exactly the same at the end of recovery as it was at the beginning of stimulation. The second and subsequent response curves are therefore identical to the first, as long as there’s been enough time for complete recovery. In this case, there’s no decrease in response intensity, meaning there’s no fatigue.
We have an exactly parallel case in muscles. ‘In muscle with normal circulation and nutrition there is always an interval between each pair of stimuli, in which the height of twitch does not diminish even after protracted excitation, and no fatigue appears.’[10]
We have an exactly parallel case in muscles. ‘In muscle with normal circulation and nutrition, there is always a gap between each pair of stimuli, during which the height of the twitch doesn’t decrease even after prolonged stimulation, and no fatigue shows up.’[10]

Fig. 20.—Record Showing Diminution of Response when Sufficient Time is not Allowed for Full Recovery
Fig. 20.—Record Showing Decrease in Response When Enough Time is Not Given for Complete Recovery
In (a) stimuli were applied at intervals of one minute; in (b) the intervals were reduced to half a minute; this caused a diminution of response. In (c) the original rhythm is restored, and the response is found to be enhanced. (Radish.)
In (a) stimuli were applied at one-minute intervals; in (b) the intervals were cut down to half a minute; this led to a decrease in response. In (c) the original rhythm is brought back, and the response is found to be improved. (Radish.)
Apparent fatigue when stimulation frequency increased.—If the rhythm of stimulation frequency be now changed, and made quicker, certain remarkable modifications will appear in the response-curves. In fig. 20, the first part shows the responses at one minute interval, by which time the individual recovery was complete.
Noticeable fatigue as the stimulation frequency increased.—If we change the stimulation frequency to a faster rhythm, we'll see some significant changes in the response curves. In fig. 20, the first part shows the responses at one-minute intervals, by which time the individual has fully recovered.
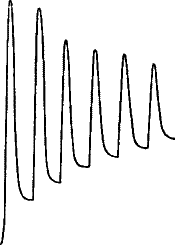
Fig. 21.—Fatigue in Celery
Fig. 21.—Tiredness in Celery
Vibration of 30° at intervals of half a minute.
Vibration of 30° every 30 seconds.
The rhythm was now changed to intervals of half a minute, instead of one, while the stimuli were maintained at the same intensity as before. It will be noticed (fig. 20, b) that these responses appear much feebler than the first set, in spite of the equality of stimulus. An inspection of the figure may perhaps throw some light on the subject. It will be seen that when greater frequency of stimulation was introduced, the tissue had not yet had time to effect complete recovery from previous strain. The molecular swing towards equilibrium had not yet abated, when the new stimulus, with its opposing impulse, was received. There is thus a diminution of height in the resultant response. The original rhythm of one minute was now restored, and the succeeding curves (fig. 20, c) at once show increased response. An analogous instance may be cited in the case of muscle response, where ‘the height of twitch diminishes more rapidly in proportion as the excitation interval is shorter.’[11]
The rhythm was now changed to intervals of half a minute instead of one, while the stimuli were kept at the same intensity as before. It will be noticed (fig. 20, b) that these responses seem much weaker than the first set, despite the stimulus being the same. Looking at the figure may shed some light on this. It shows that when a higher frequency of stimulation was introduced, the tissue hadn't fully recovered from the previous strain. The molecular swing towards equilibrium hadn't settled down yet when the new stimulus, with its opposing force, was received. As a result, there was a reduction in the height of the response. The original rhythm of one minute was restored, and the following curves (fig. 20, c) immediately show an increased response. A similar example can be seen in muscle response, where 'the height of the twitch diminishes more rapidly as the excitation interval is shorter.'[11]

Fig. 22.—Fatigue in Leaf-stalk of Cauliflower
Fig. 22.—Leaf-Stalk Fatigue in Cauliflower
Stimulus: 30° vibration at intervals of one minute.
30° vibration every minute.
From what has just been said it would appear that one of the causes of diminution of response, or fatigue, is the residual strain. This is clearly seen in fig. 21, in a record which I obtained with celery-stalk. It will be noticed there that, owing to the imperfect molecular recovery during the time allowed, the succeeding heights of the responses have undergone a continuous diminution. Fig. 22 gives a photographic record of fatigue in the leaf-stalk of cauliflower.
From what we've just discussed, it seems that one of the reasons for decreased response, or fatigue, is the leftover strain. This is clearly illustrated in fig. 21, in a record I obtained using celery stalk. You'll notice that, due to the incomplete molecular recovery during the time period allowed, the subsequent heights of the responses have continuously decreased. Fig. 22 provides a photographic record of fatigue in the leaf stalk of cauliflower.
It is evident that residual strain, other things being equal, will be greater if the stimuli have been excessive. This is well seen in fig. 23, where the set of first three curves A is for stimulus intensity of 45° vibration, and the second set B, with an augmented response, for stimulus intensity of 90° vibration. On reverting in C to stimulus intensity of 45°, the responses are seen to have undergone a great diminution as compared with the first set A. Here is seen marked fatigue, the result of overstrain from excessive stimulation.
It’s clear that residual strain, assuming everything else is the same, will be greater if the stimuli have been too intense. This is clearly illustrated in fig. 23, where the first set of three curves A corresponds to a stimulus intensity of 45° vibration, and the second set B, which shows a stronger response, corresponds to a stimulus intensity of 90° vibration. When reverting to a stimulus intensity of 45° in C, the responses show a significant decrease compared to the first set A. This indicates clear fatigue, resulting from overstrain due to excessive stimulation.

Fig. 23.—Effect of Overstrain in Producing Fatigue
Fig. 23.—How Overexertion Leads to Fatigue
Successive stimuli applied at intervals of one minute. The intensity of stimulus in C is the same as that of A, but response is feebler owing to previous over-stimulation. Fatigue is to a great extent removed after fifteen minutes’ rest, and the responses in D are stronger than those in C. The vertical line between arrows represents ·05 volt. (Turnip leaf-stalk.)
Successive stimuli were applied at one-minute intervals. The intensity of the stimulus in C is the same as in A, but the response is weaker due to earlier over-stimulation. Fatigue is mostly alleviated after fifteen minutes of rest, and the responses in D are stronger than those in C. The vertical line between arrows represents 0.05 volt. (Turnip leaf-stalk.)
If this fatigue be really due to residual strain effect, then, as strain disappears with time, we may expect the responses to regain their former height after a period of rest. In order to verify this, therefore, I renewed the stimulation (at intensity 45°) after fifteen minutes. It will at once be seen from record D how far the fatigue had been removed.
If this fatigue is really caused by leftover strain, then as the strain fades over time, we can expect the responses to return to their original levels after taking a break. To check this, I resumed the stimulation (at intensity 45°) after fifteen minutes. It will be immediately clear from record D how much the fatigue had decreased.
One peculiarity that will be noticed in these curves is that, owing to the presence of comparatively little residual strain, the first response of each set is relatively large. The succeeding responses are approximately equal where the residual strains are similar. The first response of A shows this because it had had long previous rest. The first of B shows it because we are there passing for the first time to increased stimulation. The first of C does not show it, because there is now a strong residual strain. D again shows it because the strain has been removed by fifteen minutes’ rest.
One noticeable feature in these curves is that, due to the relatively low residual strain, the initial response of each set is quite large. The following responses are roughly equal when the residual strains are similar. The first response of A demonstrates this because it had a long period of rest before. The first response of B illustrates this as we are experiencing increased stimulation for the first time. The first response of C does not show this, as there is a significant residual strain present. D shows it again because the strain has been alleviated with fifteen minutes of rest.
Fatigue under continuous stimulation.—The effect of fatigue is exhibited in marked degree when a tissue is subjected to continuous stimulation. In cases where there is marked fatigue, as for instance in certain muscles, the top of the tetanic curve undergoes rapid decline. A similar effect is obtained also with plants (fig. 24).
Fatigue under continuous stimulation.—The impact of fatigue becomes very noticeable when a tissue is exposed to continuous stimulation. In situations where fatigue is significant, such as in certain muscles, the peak of the tetanic curve quickly drops. A similar effect is also seen in plants (fig. 24).

Fig. 24.—Rapid Fatigue under Continuous Stimulation in (a) Muscle; (b) in Leaf-stalk of Celery
Fig. 24.—Quick Tiredness under Continuous Stimulation in (a) Muscle; (b) in Celery Leaf Stem
The effect of rest in producing molecular recovery, and hence in the removal of fatigue, is well illustrated in the following set of photographic records (fig. 25). The first shows the curve obtained with a fresh plant. The effect is seen to be very large. Two minutes were allowed for recovery, and then stimulation was repeated during another two minutes. The response in this case is seen to be decidedly smaller. A third case is somewhat similar to the second. A period of rest of five minutes was now allowed, and the curve obtained subsequently, owing to partial removal of residual strain, is found to exhibit greater response.
The impact of rest on molecular recovery, and thus the alleviation of fatigue, is clearly shown in the following set of photographic records (fig. 25). The first image presents the curve from a fresh plant. The effect appears to be quite significant. After a recovery period of two minutes, stimulation was repeated for another two minutes. The response in this case is noticeably smaller. A third case is somewhat similar to the second. After a rest period of five minutes, the resulting curve, due to the partial removal of residual strain, shows a greater response.

Fig. 25.—Effect of Continuous Vibration (through 50°) in Carrot
Fig. 25.—Impact of Continuous Vibration (up to 50°) on Carrot
In the first three records, two minutes’ stimulation is followed by two minutes’ recovery. The last record was taken after the specimen had a rest of five minutes. The response, owing to removal of fatigue by rest, is stronger.
In the first three records, two minutes of stimulation is followed by two minutes of recovery. The last record was taken after the specimen had a five-minute rest. The response, due to the removal of fatigue from resting, is stronger.
The results thus arrived at, under the simple conditions of vegetable life, free as they are from all possible complications and uncertainties, may perhaps throw some light on the obscure phenomena of fatigue in animal tissues.
The results obtained in these basic conditions of plant life, which are free from any possible complications and uncertainties, may help shed some light on the unclear phenomena of fatigue in animal tissues.
FOOTNOTES:
CHAPTER VI
PLANT RESPONSE—ON DIPHASIC VARIATION
- Diphasic variation
- —Positive after-effect and positive response
- —Radial E.M. variation.
When a plant is stimulated at any point, a molecular disturbance—the excitatory wave—is propagated outwards from the point of its initiation.
When a plant gets stimulated at any point, a molecular disturbance—the excitatory wave—spreads outward from where it started.
Diphasic variation.—This wave of molecular disturbance is attended by a wave of electrical disturbance. (Usually speaking, the electrical relation between disturbed and less disturbed is that of copper to zinc.) It takes some time for a disturbance to travel from one point to another, and its intensity may undergo a diminution as it recedes further from its point of origin. Suppose a disturbance originated at C; if two points are taken near each other, as A and B, the disturbance will reach them almost at the same time, and with the same intensity. The electric disturbance will be the same in both. The effect produced at A and B will balance each other and there will be no resultant current.
Diphasic variation.—This wave of molecular disturbance is accompanied by a wave of electrical disturbance. (Generally, the electrical relationship between the disturbed and the less disturbed is similar to that of copper to zinc.) It takes some time for a disturbance to move from one point to another, and its intensity may decrease as it moves further away from its origin. Imagine a disturbance starting at C; if we take two points close to each other, like A and B, the disturbance will reach them almost simultaneously and with the same intensity. The electrical disturbance will be identical at both points. The effects produced at A and B will cancel each other out, resulting in no net current.
By killing or otherwise reducing the sensibility of B as is done in the method of injury, there is no response at B, and we obtain the unbalanced response, due to disturbance at A; the same effect is obtained by putting a clamp between A and B, so that the disturbance may not reach B. But we may get response even without injury or block. If we have the contacts at A and B, and if we give a tap nearer A than B (fig. 26, a), then we have (1) the disturbance reaching A earlier than B. (2) The disturbance reaching A is much stronger than at B. The disturbance at B may be so comparatively feeble as to be negligible.
By killing or reducing the sensitivity of B as done in the injury method, there’s no response at B, resulting in an unbalanced reaction caused by the disturbance at A. We can achieve the same effect by placing a clamp between A and B, preventing the disturbance from reaching B. However, we can still get a reaction even without injury or blockage. If we have connections at A and B, and we tap closer to A than B (fig. 26, a), then (1) the disturbance reaches A before it reaches B. (2) The disturbance hitting A is significantly stronger than the one at B. The disturbance at B may be so weak that it's negligible.
It will thus be seen that we might obtain responses even without injury or block, in cases where the disturbance is enfeebled in reaching a distant point. In such a case on giving a tap near A a responsive current would be produced in one direction, and in the opposite direction when the tap is given near B (fig. 26, b). Theoretically, then, we might find a neutral point between A and B, so that, on originating the disturbance there, the waves of disturbance would reach A and B at the same instant and with the same intensity. If, further, the rate of recovery be the same for both points, then the electric disturbances produced at A and B will continue to balance each other, and the galvanometer will show no current. On taking a cylindrical root of radish I have sometimes succeeded in finding a neutral point, which, being disturbed, did not give rise to any resultant current. But disturbing a point to the right or to the left gave rise to opposite currents.
It will be clear that we can get responses even without injury or blockage, in situations where the disturbance weakens before reaching a distant point. In this case, if we tap near A, a current will respond in one direction, and if we tap near B, it will respond in the opposite direction (fig. 26, b). Theoretically, we could find a neutral point between A and B, where starting the disturbance would cause the waves to reach A and B simultaneously and with equal intensity. Furthermore, if the recovery rate is the same for both points, the electric disturbances at A and B will continue to counterbalance each other, and the galvanometer will indicate no current. When I took a cylindrical radish root, I sometimes managed to find a neutral point that, when disturbed, did not create any resultant current. However, disturbing a point to the right or left resulted in opposite currents.
It is, however, difficult to obtain an absolutely cylindrical specimen, as it always tapers in one direction. The conductivity towards the tip of the root is not exactly the same as that in the ascending direction. It is therefore difficult to fix an absolutely neutral point, but a point may be found which approaches this very nearly, and on stimulating the stalk near this, a very interesting diphasic variation has been observed. In a specimen of cauliflower-stalk, (1) stimulus was applied very much nearer A than B (the feeble disturbance reaching B was negligible). The resulting response was upward and the recovery took place in about sixty seconds.
It’s, however, challenging to obtain a perfectly cylindrical specimen, as it always tapers in one direction. The conductivity towards the tip of the root isn’t exactly the same as that in the upward direction. It is therefore tough to establish a completely neutral point, but a point can be found that comes very close, and when stimulating the stalk near this point, a fascinating diphasic variation has been observed. In a specimen of cauliflower stalk, (1) the stimulus was applied much closer to A than B (the weak disturbance that reached B was minimal). The resulting response was upward, and recovery took about sixty seconds.

(2) Stimulus was next applied near B. The resulting response was now downward (fig. 26, b).
(2) The next step was to apply the stimulus near B. The resulting response was now downward (fig. 26, b).
(3) The stimulus was now applied near the approximately neutral point N. In this case, owing to a slight difference in the rates of propagation in the two directions, a very interesting diphasic variation was produced (fig. 26, c). From the record it will be seen that the disturbance arrived earlier at A than at B. This produced an upward response. But during the subsidence of the disturbance at A, the wave reached B. The effect of this was to produce a current in the opposite direction. This apparently hastened the recovery of A (from 60 seconds to 12 seconds). The excitation of A now disappeared, and the second phase of response, that due to excitation of B, was fully displayed.
(3) The stimulus was now applied near the roughly neutral point N. In this instance, due to a slight difference in the rates of propagation in both directions, a very interesting biphasic variation was produced (fig. 26, c). From the record, you can see that the disturbance reached A before it reached B. This caused an upward response. However, during the decline of the disturbance at A, the wave finally reached B. The result of this was a current flowing in the opposite direction. This seemingly sped up the recovery of A (from 60 seconds to 12 seconds). The excitation of A then faded, and the second phase of response, which was due to the excitation of B, became fully evident.
Positive after-effect.—If we regard the response due to excitation of A as negative, the later effect on B would appear as a subsequent positive variation.
Positive after-effect.—If we see the response caused by the stimulation of A as negative, the later effect on B would be perceived as a following positive change.
In the response of nerve, for example, where contacts are made at two surfaces, injured and uninjured, there is sometimes observed, first a negative variation, and then a positive after-effect. This may sometimes at least be due to the proximal uninjured contact first giving the usual negative variation, and the more distant contact of injury giving rise, later, to the opposite, that is to say, apparently positive, response. There is always a chance of an after-effect due to this cause, unless (1) the injured end be completely killed and rendered quite irresponsive, or (2) there be an effective block between A and B, so that the disturbance due to stimulus can only act on one, and not on the other.
In nerve responses, for example, where connections are made at two surfaces, one injured and the other uninjured, it's sometimes observed that there is a negative response first, followed by a positive after-effect. This may be due to the uninjured area initially showing the typical negative response, while the injured area responds later with what appears to be a positive reaction. There’s always a possibility of an after-effect happening unless (1) the injured end is completely destroyed and becomes totally unresponsive, or (2) there is an effective barrier between A and B, so the stimulus can only influence one area and not the other.
I have found cases where, even when there was a perfect block, a positive after-effect occurred. It would thus appear that if molecular distortion from stimulus give rise to a negative variation, then during the process of molecular recovery there may be over-shooting of the equilibrium position, which may be exhibited as a positive variation.
I have found situations where, even with a complete block, a positive after-effect happened. It seems that if molecular distortion from a stimulus leads to a negative change, then during the recovery process, there might be an overshoot beyond the equilibrium position, which can show up as a positive change.
Positive variation.—The responses given by muscle or nerve are, normally speaking, negative. But that of retina is positive. The sign of response, however, is apt to be reversed if there be any molecular modification of the tissue from changes of external circumstances. Thus it is often found that nerve in a stale condition gives positive, instead of the normal negative variation, and stale retina often gives negative, instead of the usual positive.
Positive variation.—Typically, the responses from muscle or nerve are negative. However, the retina's response is positive. The nature of the response can flip if there's any molecular change in the tissue due to external factors. So, it's often observed that nerves in a stale condition show a positive response instead of the usual negative one, and stale retinas can give a negative response instead of the expected positive.

Fig. 27.—Abnormal Positive Responses in Stale Leaf-stalk of Turnip converted into Normal Negative under Strong Stimulation[12]
Fig. 27.—Unusual Positive Responses in Old Leaf-stalk of Turnip changed to Normal Negative with Strong Stimulation[12]
The relative intensities of stimuli in the two cases are in the ratio of 1:7.
The relative intensities of stimuli in the two cases are in the ratio of 1:7.
Curiously enough, I have on many occasions found exactly parallel instances in the response of plants. Plants when fresh, as stated, give negative responses as a rule. But when somewhat faded they sometimes give rise to positive response. Again, just as in the modified nerve the abnormal positive response gives place to the normal negative under strong and long-continued stimulation, so also in the modified plant the abnormal positive response passes into negative (fig. 27) under strong stimulation. I was able in some cases to trace this process of gradual reversal, by continuously increasing the intensity of stimulus. It was then found that as the stimulus was increased, the positive at a certain point underwent a reversal into the normal negative response (fig. 28).
Interestingly, I've often noticed similar reactions in plants. Fresh plants usually show negative responses. However, when they start to wilt, they sometimes begin to show positive responses. Just like in the modified nerve where an abnormal positive response changes back to the normal negative under intense and prolonged stimulation, a modified plant also shifts from an abnormal positive response to negative under strong stimulation. In some cases, I was able to observe this gradual change by steadily increasing the intensity of the stimulus. It turned out that as the stimulus increased, the positive response eventually switched back to the normal negative response at a certain point.

Fig. 28. Abnormal Positive passing into Normal Negative in a Stale Specimen of Leaf-stalk of Cauliflower
Fig. 28. Abnormal Positive transitioning into Normal Negative in an Old Sample of Cauliflower Leaf-Stalk
Stimulus was gradually increased from 1 to 10, by means of spring-tapper. When the stimulus intensity was 10, the response became reversed into normal negative. (Parts of 8 and 9 are out of the plate.)
Stimulus was gradually increased from 1 to 10, using a spring-tapper. When the stimulus intensity reached 10, the response changed to a normal negative. (Parts of 8 and 9 are out of the plate.)
The plant thus gives a reversed response under abnormal conditions of staleness. I have sometimes found similar reversal of response when the plant is subjected to the abnormal conditions of excessively high or low temperature.
The plant therefore reacts differently under unusual conditions of staleness. I have sometimes noticed a similar change in response when the plant is exposed to extreme temperatures, whether too high or too low.
Radial E.M. variation.—We have seen that a current of response flows in the plant from the relatively more to the relatively less excited. A theoretically important experiment is the following: A thick stem of plant stalk was taken and a hole bored so as to make one contact with the interior of the tissue, the other being on the surface. After a while the current of injury was found to disappear. On exciting the stem by taps or torsional vibration, a responsive current was observed which flowed inwards from the more disturbed outer surface to the shielded core inside (fig. 29). Hence it is seen that when a wave of disturbance is propagated along the plant, there is a concomitant wave of radial E.M. variation. The swaying of a tree by the wind would thus appear to give rise to a radial E.M.F.
Radial E.M. variation.—We've observed that a current of response flows in the plant from areas that are more excited to those that are less excited. An important theoretical experiment is as follows: A thick stem of a plant was taken, and a hole was drilled to connect the interior tissue to the surface. After some time, the injury current was found to disappear. When the stem was stimulated by taps or twisting vibrations, a responsive current was noted flowing inward from the more disturbed outer surface to the protected core inside (fig. 29). Therefore, it is evident that when a wave of disturbance travels along the plant, there is a corresponding wave of radial E.M. variation. The swaying of a tree in the wind would thus seem to generate a radial E.M.F.

FOOTNOTES:
[12] For general purposes it is immaterial whether the responses are recorded up or down. For convenience of inspection they are in general recorded up. But in cases where it is necessary to discriminate the sign of response, positive response will be recorded up, and negative down.
[12] For general purposes, it doesn’t really matter if the responses are recorded as positive or negative. For easier review, they are usually recorded up. However, in situations where it's important to specify the nature of the response, a positive response will be noted as up, and a negative response as down.
CHAPTER VII
PLANT RESPONSE—ON THE RELATION BETWEEN STIMULUS AND RESPONSE
- Increased response with increasing stimulus
- —Apparent diminution of response with excessively strong stimulus.
As already said, in the living tissue, molecular disturbance induced by stimulus is accompanied by an electric disturbance, which gradually disappears with the return of the disturbed molecules to their position of equilibrium. The greater the molecular distortion produced by the stimulus, the greater is the electric variation produced. The electric response is thus an outward expression of a molecular disturbance produced by an external agency, the stimulus.
As mentioned earlier, in living tissue, the molecular disruption caused by a stimulus is accompanied by an electrical disturbance, which gradually fades as the affected molecules return to their equilibrium state. The more significant the molecular distortion caused by the stimulus, the larger the electrical variation produced. Therefore, the electrical response is an outward manifestation of the molecular disruption caused by an external factor, the stimulus.
Curve of relation between stimulus and response.—In the curve showing the relation between stimulus and response in nerve and muscle, it is found that the molecular effect as exhibited either by contraction or E.M. variation in muscle, or simply by E.M. variation in nerve, is at first slight. In the second part, there is a rapidly increasing effect with increased stimulus. Finally, a tendency shows itself to approach a limit of response. Thus we find the curve at first slightly convex, then straight and ascending, and lastly, concave to the abscissa (fig. 30).
Curve of relationship between stimulus and response.—In the curve illustrating the relationship between stimulus and response in nerves and muscles, we observe that the molecular effect, whether shown by contraction or E.M. variation in muscles, or simply by E.M. variation in nerves, starts off as minimal. In the next phase, there is a rapid increase in effect with a larger stimulus. Eventually, there seems to be a limit to the response. Therefore, we see the curve starting off slightly convex, then straight and rising, and finally concave to the horizontal axis (fig. 30).
In muscle the limit of response is reached much sooner than in nerve. As will be seen, the range of variation of stimulus in these curves is not very great. When the stimulus is carried beyond moderate limits, the response, owing to fatigue or other causes, may sometimes undergo an actual diminution.

Fig. 30.—Curves Showing the Relation Between the Intensity of Stimulus and Response
Fig. 30.—Curves Showing the Relationship Between the Intensity of the Stimulus and the Response
Abscissæ indicate increasing intensity of stimulus. Ordinates indicate magnitude of response. (Waller.)
Abscissae show the increasing strength of the stimulus. Ordinates indicate the level of response. (Waller.)

Fig. 31
Fig. 31
Taps of increasing strength 1:2:3:4 producing increased response in leaf stalk of turnip.
Taps with increasing strength 1:2:3:4 lead to a stronger response in the leaf stalk of the turnip.
I have obtained very interesting results, with reference to the relation between stimulus and response, when experimenting with plants. These results are suggestive of various types of response met with in animal tissues.
I have gotten some very interesting results regarding the relationship between stimulus and response while experimenting with plants. These results hint at different types of responses seen in animal tissues.
1. In order to obtain the simplest type of effects, not complicated by secondary phenomena, one has to choose specimens which exhibit little fatigue. Having procured these, I undertook two series of experiments. In the first (A) the stimulus was applied by means of the spring-tapper, and in the second (B) by torsional vibration.
1. To get the simplest type of effects, free from secondary phenomena, you need to choose samples that show little fatigue. After acquiring these, I conducted two sets of experiments. In the first (A), the stimulus was applied using a spring-tapper, and in the second (B), through torsional vibration.
(A) The first stimulus was given by a fall of the lever through h, the second through 2 h, and so on. The response-curves clearly show increasing effect with increased stimulus (fig. 31).
(A) The first stimulus was triggered by the lever dropping by h, the second by 2 h, and so on. The response curves clearly demonstrate a stronger effect with a greater stimulus (fig. 31).
Fig. 32.—Increased Response with Increasing Vibrational Stimuli (Cauliflower-stalk)
Fig. 32.—Higher Response with More Vibrational Stimuli (Cauliflower-stalk)
Stimuli applied at intervals of three minutes. Vertical line = ·1 volt.
Stimuli were applied every three minutes. Vertical line = ·1 volt.
(B) 1. The vibrational stimulus was increased from 2·5° to 5° to 7·5° to 10° to 12·5° in amplitude. It will be observed how the intensity of response tends to approach a limit (fig. 32).
(B) 1. The vibrational stimulus was raised from 2.5° to 5° to 7.5° to 10° to 12.5° in amplitude. You’ll notice how the intensity of the response starts to reach a limit (fig. 32).
Table showing the Increased E.M. Variation
produced by Increasing Stimulus
| Angle of Vibration | E.M.F |
|---|---|
| 2·5° | ·044 volt |
| 5° | ·075 volt |
| 7·5° | ·090 volt |
| 10° | ·100 volt |
| 12·5° | ·106 volt |
2. The next figure shows how little variation is produced with low value of stimulus, but with increasing stimulus the response undergoes a rapid increase, after which it tends to approach a limit (fig. 33, a).
2. The next figure illustrates how minimal change occurs with a low level of stimulus, but as the stimulus increases, the response quickly escalates, eventually approaching a limit (fig. 33, a).

Fig. 33.—Responses to Increasing Stimuli produced by Increasing Angle of Vibration
Fig. 33.—Responses to Increasing Stimuli caused by Increasing Angle of Vibration
(a) Record with a specimen of fresh radish. Stimuli applied at intervals of two minutes. The record is taken for one minute.
(a) Record with a sample of fresh radish. Stimuli applied every two minutes. The record is taken for one minute.
(b) Record for stale radish. There is a reversed response for the feeble stimulus of 5° vibration.
(b) Record for stale radish. There is an opposite reaction for the weak stimulus of 5° vibration.
3. As an extreme instance of the case just cited, I have often come across a curious phenomenon. During the gradual increase of the stimulus from a low value there would be apparently no response. But when a critical value was reached a maximum response would suddenly occur, and would not be exceeded when the stimulus was further increased. Here we have a parallel to what is known in animal physiology as the ‘all or none’ principle. With the cardiac muscle, for example, there is a certain minimal intensity which is effective in producing response, but further increase of stimulus produces no increase in response.
3. As an extreme example of the case just mentioned, I've often come across a curious phenomenon. During the gradual increase of the stimulus from a low level, there seems to be no reaction. However, when a certain critical level is reached, a maximum response suddenly occurs and won’t increase even if the stimulus is raised further. This is similar to what is known in animal physiology as the ‘all or none’ principle. For instance, with cardiac muscle, there’s a specific minimal intensity that effectively produces a response, but increasing the stimulus further doesn’t result in a greater response.
4. From an inspection of the records of responses which are given, it will be seen that the slope of a curve which shows the relation of stimulus to response will at first be slight, the curve will then ascend rapidly, and at high values of stimulus tend to become horizontal. The curve as a whole becomes, first slightly convex to the abscissa, then straight and ascending, and lastly concave. A far more pronounced convexity in the first part is shown in some cases, especially when the specimen is stale. This is due to the fact that under these circumstances response is apt to begin with an actual reversal of sign, the plant under feebler than a certain critical intensity of stimulus giving positive, instead of the normal negative, response (fig. 33, b).
4. Looking at the response records , you will notice that the slope of the curve showing the relationship between stimulus and response starts off gradually, then quickly rises, and at high stimulus levels tends to level off. Overall, the curve initially appears slightly convex to the horizontal axis, then becomes straight and rising, and finally curves downwards. In some cases, especially when the specimen is old, the initial part shows a more pronounced convexity. This happens because, under these conditions, the response may begin with an actual reversal in sign, where the plant responds positively rather than the usual negative response when the stimulus is below a certain critical intensity (fig. 33, b).
Diminution of response with excessively strong stimulus.—It is found that in animal tissues there is sometimes an actual diminution of response with excessive increase of stimulus. Thus Waller finds, in working with retina, that as the intensity of light stimulus is gradually increased, the response at first increases, and then sometimes undergoes a diminution. This phenomenon is unfortunately complicated by fatigue, itself regarded as obscure. It is therefore difficult to say whether the diminution of response is due to fatigue or to some reversing action of an excessively strong stimulus.
Decrease in response with overly strong stimulus.—It has been observed that in animal tissues, there can be an actual decrease in response when the stimulus becomes too intense. For example, Waller found that while working with the retina, as the intensity of light stimulus is gradually increased, the response initially goes up but then sometimes decreases. This phenomenon is complicated by fatigue, which is also not fully understood. Because of this, it's hard to determine whether the decrease in response is caused by fatigue or by some reversing effect of an overly strong stimulus.
From fig. 33, b, above, it is seen that there was an actual reversal of response in the lower portion of the curve. It is therefore not improbable that there may be more than one point of reversal.
From fig. 33, b, above, it can be observed that there was a genuine reversal of response in the lower part of the curve. It’s therefore quite possible that there may be more than one point of reversal.
In physical phenomena we are, however, acquainted with numerous instances of reversals. For example, a common effect of magnetisation is to produce an elongation of an iron rod. But Bidwell finds that as the magnetising force is pushed to an extreme, at a certain point elongation ceases and is succeeded, with further increase of magnetising force, by an actual contraction. Again a photographic plate, when exposed continuously to light, gives at first a negative image. Still longer exposure produces a positive. Then again we have a negative. There is thus produced a series of recurrent reversals. In photographic prints of flashes of lightning, two kinds of images are observed, one, the positive—when the lightning discharge is moderately intense—and the other, negative, the so-called ‘dark lightning’—due to the reversal action of an intensely strong discharge.
In physical phenomena, we often see many examples of reversals. For instance, a common effect of magnetization is that it causes an iron rod to stretch. However, Bidwell discovers that as the magnetizing force increases to an extreme level, there comes a point where the stretching stops and is followed, with further increases in the magnetizing force, by actual contraction. Similarly, when a photographic plate is continuously exposed to light, it initially produces a negative image. Longer exposure leads to a positive image, and then it returns to a negative once again. This creates a series of recurring reversals. In photographic prints of lightning strikes, two types of images are seen: one positive, which occurs when the lightning discharge is of moderate intensity, and the other negative, known as 'dark lightning,' which results from the reversing action of a very strong discharge.
In studying the changes of conductivity produced in metallic particles by the stimulus of Hertzian radiation, I have often noticed that whereas feeble radiation produces one effect, strong radiation produces the opposite. Again, under the continuous action of electric radiation, I have frequently found recurrent reversals.[13]
In studying how conductivity changes in metal particles due to Hertzian radiation, I've often observed that weak radiation causes one effect, while strong radiation causes the opposite. Additionally, with the ongoing influence of electric radiation, I've frequently noticed recurring reversals.[13]
Diminution of response under strong stimulus traced to fatigue.—But there are instances in plant response where the diminution effect can be definitely traced to fatigue. The records of these cases are extremely suggestive as to the manner in which the diminution is brought about. The accompanying figures (fig. 34) give records of responses to increasing stimulus. They were made with specimens of cauliflower-stalks, one of which (a) showed little fatigue, while in the other (b) fatigue was present. It will be seen that the curves obtained by joining the apices of the successive single responses are very similar.
Decreased response under strong stimulus linked to fatigue.—However, there are cases in plant responses where the decrease can be clearly linked to fatigue. The records of these instances are very informative regarding how this decrease occurs. The figures (fig. 34) provide data on responses to increasing stimulus. These were gathered using cauliflower stalk specimens, one of which (a) showed minimal fatigue, while in the other (b) fatigue was evident. It can be observed that the curves created by connecting the peaks of the successive individual responses are quite similar.

Fig. 34.—Responses to Increasing Stimulus obtained with Two Specimens of Stalk of Cauliflower
Fig. 34.—Responses to Increasing Stimulus obtained with Two Specimens of Cauliflower Stalk
In (a) fatigue is absent, in (b) it is present.
In (a) there is no fatigue, in (b) it is present.
In one case there is no fatigue, the recovery from each stimulus being complete. Every response in the series therefore starts from a position of perfect equilibrium, and the height of the single responses increases with increasing stimulation. But in the second case, the strain is not completely removed after any single stimulation of the series. That recovery is partial is seen by the gradual shifting of the base line upwards. In the former case the base line is horizontal and represents a condition of complete equilibrium. Now, however, the base line, or line of modified equilibrium, is tilted upwards. Thus even in this case if we measure the heights of successive responses from the line of absolute equilibrium, they will be found to increase with increasing stimulus. Ordinarily, however, we make no allowance for the shifting of the base line, measuring response rather from the place of its previous recovery, or from the point of modified equilibrium. Judged in this way, the responses undergo an apparent diminution.
In one scenario, there’s no fatigue, and recovery from each stimulus is complete. So, every response in the series begins at a point of perfect balance, and the intensity of individual responses grows with stronger stimulation. But in the second scenario, the strain isn’t fully removed after any single stimulation in the series. The fact that recovery is only partial is shown by the gradual rise of the base line. In the first case, the base line is flat and indicates total equilibrium. Now, however, the base line, or line of altered equilibrium, is slanted upwards. Thus, even in this situation, if we measure the heights of successive responses from the line of perfect equilibrium, we’ll find that they increase with stronger stimuli. Typically, though, we don’t account for the shift in the base line and instead measure responses based on their previous recovery point or from the modified equilibrium point. When assessed this way, the responses seem to decrease.
FOOTNOTES:
CHAPTER VIII
PLANT RESPONSE—ON THE INFLUENCE OF TEMPERATURE
- Effect of very low temperature
- —Influence of high temperature
- —Determination of death-point
- —Increased response as after-effect of temperature variation
- —Death of plant and abolition of response by the action of steam.
For every plant there is a range of temperature most favourable to its vital activity. Above this optimum, the vital activity diminishes, till a maximum is reached, when it ceases altogether, and if this point be maintained for a long time the plant is apt to be killed. Similarly, the vital activity is diminished if the temperature be lowered below the optimum, and again, at a minimum point it ceases, while below this minimum the plant may be killed. We may regard these maximum and minimum temperatures as the death-points. Some plants can resist these extremes better than others. Length of exposure, it should however be remembered, is also a determining factor in the question as to whether or not the plant shall survive unfavourable conditions of temperature. Thus we have hardy plants, and plants that are affected by excessive variations of temperature. Within the characteristic power of the species, there may be, again, a certain amount of individual difference.
For every plant, there's a specific temperature range that's best for its growth. When the temperature goes above this ideal range, the plant's activity decreases until it reaches a maximum point, where it stops completely. If this high temperature is sustained for too long, the plant can die. Likewise, if the temperature drops below the ideal range, the plant's activity also decreases, and at a certain low point, it stops entirely; below this minimum, the plant may also die. We can think of these maximum and minimum temperatures as the points of death. Some plants can handle these extremes better than others. However, the length of time a plant is exposed to these unfavorable temperatures is also an important factor in determining whether it will survive. So, we have hardy plants and those that are more sensitive to significant temperature changes. Within each species, there can also be individual differences in how they cope with these conditions.
These facts being known, I was anxious to deter mine whether the undoubted changes induced by temperature in the vital activity of plants would affect electrical response.
These facts known, I was eager to determine whether the clear changes caused by temperature in the vital activity of plants would impact electrical response.
Effect of very low temperature.—As regards the influence of very low temperature, I had opportunities of studying the question on the sudden appearance of frost. In the previous week, when the temperature was about 10° C., I had obtained strong electric response in radishes whose value varied from ·05 to ·1 volt. But two or three days later, as the effect of the frost, I found electric response to have practically disappeared. A few radishes were, however, found somewhat resistant, but the electric response had, even in these cases, fallen from the average value of ·075 V. under normal temperature to ·003 V. after the frost. That is to say, the average sensitiveness had been reduced to about 1/25th. On warming the frost-bitten radish to 20° C. there was an appreciable revival, as shown by increase in response. In specimens where the effect of frost had been very great, i.e. in those which showed little or no electric response, warming did not restore responsiveness. From this it would appear that frost killed some, which could not be subsequently revived, whereas others were only reduced to a condition of torpidity, from which there was revival on warming.
Effect of very low temperature.—Regarding the impact of very low temperatures, I had the chance to explore this issue when frost appeared suddenly. The previous week, when the temperature was around 10° C., I recorded strong electrical responses in radishes, ranging from 0.05 to 0.1 volts. However, two or three days later, following the onset of frost, I discovered that the electrical response had nearly vanished. A few radishes remained somewhat resilient, but even in these cases, the electrical response dropped from the average value of 0.075 V at normal temperatures to 0.003 V after the frost. This means the average sensitivity was reduced to about 1/25th. When I warmed the frost-damaged radish to 20° C., there was a noticeable revival, reflected by an increase in response. In specimens severely affected by frost, meaning those with little or no electrical response, warming did not restore their responsiveness. It seems that frost killed some radishes, which could not be revived later, while others only experienced a state of inactivity, from which they could recover when warmed.

Fig. 35.—Diminution of Response in Eucharis by Lowering of Temperature
Fig. 35.—Reduced Response in Eucharis by Decreasing Temperature
(a) Normal response at 17° C.
(b) The response almost disappears when plant is subjected to −2° C.
for fifteen minutes.
(c) Revival of response on warming to 20° C.
(a) Normal response at 17° C.
(b) The response nearly vanishes when the plant is exposed to -2° C for fifteen minutes.
(c) Response returns upon warming to 20° C.
I now tried the effect of artificial lowering of temperature on various plants. A plant which is very easily affected by cold is a certain species of Eucharis lily. I first obtained responses with the leaf-stalk of this lily at the ordinary temperature of the room (17° C.). I then placed it for fifteen minutes in a cooling chamber, temperature −2° C., for only ten minutes, after which, on trying to obtain response, it was found to have practically disappeared. I now warmed the plant by immersing it for awhile in water at 20° C., and this produced a revival of the response (fig. 35). If the plant be subjected to low temperature for too long a time, there is then no subsequent revival.
I now tested how artificially lowering the temperature affects various plants. One plant that is really sensitive to cold is a specific type of Eucharis lily. I first observed its response at room temperature (17° C.). Then, I placed it in a cooling chamber at −2° C. for fifteen minutes. After just ten minutes, when I tried to get a response, it was almost gone. I then warmed the plant by soaking it in water at 20° C., which revived its response (fig. 35). However, if the plant is exposed to low temperatures for too long, it won’t recover afterward.
I obtained a similar marked diminution of response with the flower-stalk of Arum lily, on lowering the temperature to zero.
I noticed a similar significant decrease in response with the flower stem of the Arum lily when I dropped the temperature to zero.
My next attempt was to compare the sensibility of different plants to the effect of lowered temperatures. For this purpose I chose three specimens: (1) Eucharis lily; (2) Ivy; and (3) Holly. I took their normal response at 17° C., and found that, generally speaking, they attained a fairly constant value after the third or fourth response. After taking these records of normal response, I placed the specimens in an ice-chamber, temperature 0° C., for twenty-four hours, and afterwards took their records once more at the ordinary temperature of the room. From these it will be seen that while the responsiveness of Eucharis lily, known to be susceptible to the effect of cold, had entirely disappeared, that of the hardier plants, Holly and Ivy, showed very little change (fig. 36).
My next attempt was to compare how different plants reacted to lower temperatures. For this, I chose three specimens: (1) Eucharis lily; (2) Ivy; and (3) Holly. I recorded their normal response at 17° C and found that, generally, they reached a fairly consistent value after the third or fourth response. After taking these normal response records, I placed the specimens in an ice chamber, at a temperature of 0° C, for twenty-four hours, and then recorded their responses again at the normal room temperature. From these records, it was evident that while the responsiveness of the Eucharis lily, which is known to be sensitive to cold, had completely disappeared, the hardier plants, Holly and Ivy, showed very little change (fig. 36).
Another very curious effect that I have noticed is that when a plant approaches its death-point by reason of excessively high or low temperature, not only is its general responsiveness diminished almost to zero, but even the slight response occasionally becomes reversed.
Another very curious effect that I've noticed is that when a plant is nearing its death due to extremely high or low temperatures, not only does its overall responsiveness drop almost to zero, but even the minimal responses it might have occasionally become reversed.

Fig. 36.—After-effect of Cold on Ivy, Holly, and Eucharis Lily
Fig. 36.—After-effect of Cold on Ivy, Holly, and Eucharis Lily
a. The normal response; b. Response after subjection to freezing temperature for twenty-four hours.
a. The normal response; b. Response after being exposed to freezing temperatures for twenty-four hours.
Influence of high temperature, and determination of death-point.—I next tried to find out whether a rise of temperature produced a depression of response, and whether the response disappeared at a maximum temperature—the temperature of death-point. For this purpose I took a batch of six radishes and obtained from them responses at gradually increasing temperatures. These specimens were obtained late in the season, and their electric responsiveness was much lower than those obtained earlier. The plant, previously kept for five minutes in water at a definite temperature (say 17° C.), was mounted in the vibration apparatus and responses observed. The plant was then dismounted, and replaced in the water-bath at a higher temperature (say 30° C.) again, for five minutes. A second set of responses was now taken. In this way observations were made with each specimen till the temperature at which response almost or altogether ceased was reached. I give below a table of results obtained with six specimens of radish, from which it would appear that response begins to be abolished in these cases at temperatures varying from 53° to 55° C.
Influence of high temperature and determination of death-point.—Next, I wanted to find out if an increase in temperature caused a decrease in response and if the response disappeared at a maximum temperature—the temperature of death-point. For this, I took a batch of six radishes and recorded their responses at gradually increasing temperatures. These samples were collected late in the season, and their electric responsiveness was much lower than those collected earlier. The plant, previously kept for five minutes in water at a specific temperature (let's say 17° C.), was set up in the vibration apparatus and responses were observed. The plant was then removed and placed back in the water-bath at a higher temperature (about 30° C.) for another five minutes. A second set of responses was then taken. Observations were made with each specimen until we reached the temperature at which the response nearly or completely stopped. Below is a table of results obtained from the six radish specimens, indicating that the response begins to be eliminated at temperatures ranging from 53° to 55° C.
Table showing Effect of High Temperature in Abolishing Response
| Temperature | Galvanometric response (100 dns. = ·07 V.) |
||
|---|---|---|---|
| (1) I'm ready for the text. Please provide it. | 17° C | 70 | dns. |
| 53° | 4 | " | |
| (2) I'm ready for the text. | 17° | 160 | " |
| 53° | 1 | " | |
| (3) Please provide the text you would like me to modernize. | 17° | 100 | " |
| 50° | 2 | " | |
| (4) { | 17° | 80 | " |
| 55° | 0 | " | |
| (5) { | 17° | 40 | " |
| 60° | 0 | " | |
| (6) Please provide the text you would like me to modernize. | 17° | 60 | " |
| 55° | 0 | " | |

Fig. 37.—The Glass Chamber containing the Plant
Fig. 37.—The Glass Chamber holding the Plant
Amplitude of vibration which determines the intensity of stimulus is measured by the graduated circle seen to the right. Temperature is regulated by the electric heating coil R. For experiments on action of anæsthetics, vapour of chloroform is blown in through the side tube.
Amplitude of vibration, which determines how strong the stimulus is, is measured by the graduated circle on the right. The temperature is controlled by the electric heating coil R. For experiments on the effects of anesthetics, chloroform vapor is directed in through the side tube.
Electric heating.—The experiments just described were, however, rather troublesome, inasmuch as, in order to produce each variation of temperature, the specimen had to be taken out of the apparatus, warmed, and remounted. I therefore introduced a modification by which this difficulty was obviated. The specimen was now enclosed in a glass chamber (fig. 37), which also contained a spiral of German-silver wire, through which electric currents could be sent, for the purpose of heating the chamber. By varying the intensity of the current, the temperature could be regulated at will. The specimen chosen for experiment was the leaf-stalk of celery. It was kept at each given temperature for ten minutes, and two records were taken during that time. It was then raised by 10° C., and the same process was repeated. It will be noticed from the record (fig. 38) that in this particular case, as the temperature rose from 20° C. to 30° C., there was a marked diminution of response. At the same time, in this case at least, recovery was quicker. At 20° C., for example, the response was 21 dns., and the recovery was not complete in the course of a minute. At 30° C., however, the response had been reduced to 7·5 divisions, but there was almost complete recovery in twelve seconds. As the temperature was gradually increased, a continuous decrease of response occurred. This diminution of response with increased temperature appears to be universal, but the quickening of recovery may be true of individual cases only.
Electric heating.—The experiments I just described were pretty annoying because to change the temperature, I had to remove the sample from the apparatus, heat it, and put it back again. So, I came up with a solution to make this easier. The sample was placed in a glass chamber (fig. 37), which also had a coil of German-silver wire that could carry electric currents to heat the chamber. By adjusting the current's intensity, I could control the temperature as needed. The sample I chose for the experiment was the leaf-stalk of celery. It was kept at each specified temperature for ten minutes, during which I took two recordings. Then, the temperature was increased by 10° C., and I repeated the process. You can see from the record (fig. 38) that in this case, as the temperature went from 20° C. to 30° C., there was a noticeable decrease in response. At the same time, recovery was faster in this instance. At 20° C., for example, the response was 21 dns., and recovery wasn't complete within a minute. However, at 30° C., the response dropped to 7.5 divisions, but recovery was almost complete in twelve seconds. As I gradually increased the temperature, there was a consistent decrease in response. This decrease in response with higher temperatures seems to be a general trend, but the speed of recovery might vary in individual cases.

Fig. 38.—Effect of Temperature on Response
Fig. 38.—Impact of Temperature on Reaction
The response was abolished at the hot-water temperature of 55° C.
The response was eliminated at a hot-water temperature of 55° C.
Table showing Diminution of Response with Increasing Temperature
(·01 Volt = 35 divisions)
(·01 Volt = 35 divisions)
| Temperature | Response |
|---|---|
| 20° | 21 |
| 30° | 7·5 |
| 40° | 5·5 |
| 50° | 4 |
| 65° | 3 |
In radishes response disappeared completely at 55° C., but with celery, heated in the manner described, I could not obtain its entire abolition at 60° C. or even higher. A noticeable circumstance, however, was the prolongation of the period of recovery at these high temperatures. I soon understood the reason of this apparent anomaly. The method adopted in the present case was that of dry heating, whereas the previous experiments had been carried on by the use of hot water. It is well known that one can stand a temperature of 100° C. without ill effects in the hot-air chamber of a Turkish bath, while immersion in water at 100° C. would be fatal.
In radishes, the response completely disappeared at 55°C, but with celery, when heated as described, I couldn’t achieve total loss of response at 60°C or even higher. However, a noticeable factor was the extended recovery time at these high temperatures. I soon figured out the reason for this seeming discrepancy. The method used here was dry heating, whereas the earlier experiments had utilized hot water. It’s well known that one can tolerate a temperature of 100°C without negative effects in the hot-air chamber of a Turkish bath, while immersion in water at 100°C would be deadly.
In order to find out whether subjection to hot water would kill the celery-stalk, I took it out and placed it for five minutes in water at 55° C. This, as will be seen from the record taken afterwards, effectively killed the plant (fig. 38, w).
To find out if hot water would kill the celery stalk, I took it out and put it for five minutes in water at 55° C. This, as will be evident from the record taken later, effectively killed the plant (fig. 38, w).

Fig. 39.—Effect of Rising and Falling Temperature on the Response Of Scotch Kale
Fig. 39.—Impact of Increasing and Decreasing Temperature on the Response of Scotch Kale
Increased sensitiveness as after-effect of temperature variation.—A very curious effect of temperature variation is the marked increase of sensitiveness which often appears as its after-effect. I noticed this first in a series of observations where records were taken during the rise of temperature and continued while the temperature was falling (fig. 39). The temperature was adjusted by electric heating. It was found that the responses were markedly enhanced during cooling, as compared with responses given at the same temperatures while warming (see table). Temperature variation thus seems to have a stimulating effect on response, by increasing molecular mobility in some way. The second record (fig. 40) shows the variation of response in Eucharis lily (1) during the rise, and (2) during the fall of temperature. Fig. 41 gives a curve of variation of response during the rise and fall of temperature.
Increased sensitivity as an after-effect of temperature change.—A very interesting effect of temperature change is the significant increase in sensitivity that often occurs afterward. I first noticed this in a series of observations where records were taken during the temperature rise and continued as it was falling (fig. 39). The temperature was adjusted using electric heating. It was observed that the responses were significantly heightened during cooling, as compared to responses at the same temperatures while warming (see table). Thus, temperature variation appears to have a stimulating effect on response by increasing molecular movement in some way. The second record (fig. 40) shows the variation in response of the Eucharis lily (1) during the rise and (2) during the fall of temperature. Fig. 41 provides a curve showing the variation of response during the rise and fall of temperature.
Table showing the Variation of Response in Scotch Kale
during the Rise and Fall of Temperature
| Temperature | ↓ | Response [Temperature falling] |
Response [Temperature rising] |
↑ | |||
|---|---|---|---|---|---|---|---|
| 19° C. | 47 | dns. | — | ||||
| 25° " | 24 | " | — | ||||
| 30° " | 11 | " | 23 | dns. | |||
| 50° " | 8 | " | 16 | " | |||
| 70° " | 7 | " | — | ||||
| → | |||||||

Fig. 40.—Records of Responses in Eucharis Lily during Rise and Fall of Temperature
Fig. 40.—Records of Responses in Eucharis Lily during Rise and Fall of Temperature
Stimulus constant, applied at intervals of one minute. The temperature of plant-chamber gradually rose on starting current in the heating coil; on breaking current, the temperature fell gradually. Temperature corresponding to each record is given below.
Stimulus constant, applied at one-minute intervals. The temperature of the plant chamber gradually increased when the current was turned on in the heating coil; when the current was turned off, the temperature gradually decreased. The temperature corresponding to each record is provided below.
Temperature rising: (1) 20°, (2) 20°, (3) 22°, (4) 38°, (5) 53°, (6) 68°, (7) 65°.
Temperature rising: (1) 20°, (2) 20°, (3) 22°, (4) 38°, (5) 53°, (6) 68°, (7) 65°.
Temperature falling: (8) 60°, (9) 51°, (10) 45°, (11) 40°, (12) 38°.
Temperature falling: (8) 60°, (9) 51°, (10) 45°, (11) 40°, (12) 38°.
Point of temperature maximum.—We have seen how, in cases of lowered temperature, response is abolished earlier in plants like Eucharis, which are affected by cold, than in the hardier plants such as Holly and Ivy. Plants again are unequally affected as regards the upper range. In the case of Scotch kale, for instance, response disappears after ten minutes of water temperature of about 55° C., but with Eucharis fairly marked response can still be obtained after such immersion and does not disappear till it has been subjected for ten minutes to hot water, at a temperature of 65° C. or even higher. The reason of this great power of resistance to heat is probably found in the fact that the Eucharis is a tropical plant, and is grown, in this country, in hot-houses where a comparatively high temperature is maintained.
Point of temperature maximum.—We've seen that in cases of lower temperatures, plants like Eucharis, which are sensitive to cold, lose their responsiveness sooner than hardier plants like Holly and Ivy. Similarly, plants are affected differently when it comes to higher temperatures. For example, Scotch kale stops responding after ten minutes at a water temperature of around 55° C., while Eucharis can still show a noticeable response after that time and doesn't lose it until it's been in hot water at 65° C. or even higher for ten minutes. This strong resistance to heat is likely due to the fact that Eucharis is a tropical plant, typically grown in hot-houses in this country where relatively high temperatures are kept.

Fig. 41.—Curve showing Variation of Response in Eucharis with the Rise and Fall of Temperature
Fig. 41.—Graph showing Changes in Response in Eucharis with Temperature Fluctuations
The effect of steam.—I next wished to obtain a continuous record by which the effects of suddenly increased temperatures, culminating in the death of the plant, might be made evident. For this purpose I mounted the plant in the glass chamber, into which steam could be introduced. I had chosen a specimen which gave regular response. On the introduction of steam, with the consequent sudden increase of temperature, there was a transitory augmentation of excitability. But this quickly disappeared, and in five minutes the plant was effectively killed, as will be seen graphically illustrated in the record (fig. 42).
The effect of steam.—Next, I wanted to create a continuous record that would show how sudden increases in temperature, leading to the death of the plant, could be observed. To do this, I placed the plant in a glass chamber where steam could be introduced. I selected a specimen that responded consistently. When I introduced the steam and the temperature increased suddenly, there was a brief spike in excitability. However, this quickly faded, and within five minutes, the plant was effectively dead, as will be clearly shown in the record (fig. 42).

Fig. 42.—Effect of Steam in Killing Response
Fig. 42.—Impact of Steam on Killing Response
The two records to the left exhibit normal response at 17° C. Sudden warming by steam produced at first an increase of response, but five minutes exposure to steam killed the plant (carrot) and abolished the response.
The two records on the left show a normal response at 17° C. A sudden increase in temperature from steam first caused the response to rise, but after five minutes of exposure to the steam, it killed the plant (carrot) and ended the response.
Vibrational stimulus of 30° applied at intervals of one minute; vertical line = ·1 volt.
Vibrational stimulus of 30° applied every minute; vertical line = ·1 volt.
It will thus be seen that those modifications of vital activity which are produced in plants by temperature variation can be very accurately gauged by electric response. Indeed it may be said that there is no other method by which the moment of cessation of vitality can be so satisfactorily distinguished. Ordinarily, we are able to judge that a plant has died, only after various indirect effects of death, such as withering, have begun to appear. But in the electric response we have an immediate indication of the arrest of vitality, and we are thereby enabled to determine the death-point, which it is impossible to do by any other means.
It’s clear that changes in plant activity caused by temperature variations can be accurately measured by their electric response. In fact, it can be said that there’s no other method that can clearly identify when a plant's vitality has stopped. Usually, we only realize a plant has died after it shows various signs like wilting. However, the electric response gives us an immediate sign of when vitality has halted, allowing us to pinpoint the exact death point, which isn’t possible through any other method.
It may be mentioned here that the explanation suggested by Kunkel, of the response being due to movement of water in the plant, is inadequate. For in that case we should expect a definite stimulation to be under all conditions followed by a definite electric response, whose intensity and sign should remain invariable. But we find, instead, the response to be profoundly modified by any influence which affects the vitality of the plant. For instance, the response is at its maximum at an optimum temperature, a rise of a few degrees producing a profound depression; the response disappears at the maximum and minimum temperatures, and is revived when brought back to the optimum. Anæsthetics and poisons abolish the response. Again, we have the response undergoing an actual reversal when the tissue is stale. All these facts show that mere movement of water could not be the effective cause of plant response.
It should be noted that Kunkel's explanation of the response being caused by the movement of water in the plant is insufficient. In that case, we would expect a consistent stimulation to produce a reliable electric response, with both its intensity and sign staying the same. However, we actually see that the response is greatly influenced by anything that affects the plant's vitality. For example, the response is strongest at an optimal temperature; just a few degrees increase causes a significant drop. The response disappears at both the highest and lowest temperatures but comes back when the temperature is returned to optimal. Anesthetics and toxins eliminate the response. Furthermore, the response can even reverse when the tissue is no longer fresh. All these observations indicate that just the movement of water cannot be the main reason for plant responses.
CHAPTER IX
PLANT RESPONSE—EFFECT OF ANÆSTHETICS AND POISONS
- Effect of anæsthetics, a test of vital character of response
- —Effect of chloroform
- —Effect of chloral
- —Effect of formalin
- —Method in which response is unaffected by variation of resistance
- —Advantage of block method
- —Effect of dose.
The most important test by which vital phenomena are distinguished is the influence on response of narcotics and poisons. For example, a nerve when narcotised by chloroform exhibits a diminishing response as the action of the anæsthetic proceeds. (See below, fig. 43.) Similarly, various poisons have the effect of permanently abolishing all response. Thus a nerve is killed by strong alkalis and strong acids. I have already shown how plants which previously gave strong response did not, after application of an anæsthetic or poison, give any response at all. In these cases it was the last stage only that could be observed. But it appeared important to be able to trace the growing effect of anæsthetisation or poisoning throughout the process. There were, however, two conditions which it at first appeared difficult to meet. First it was necessary to find a specimen which would normally exhibit no fatigue, and give rise for a long time to a uniform series of response. The immediate changes made in the response, in consequence of the application of chemical reagents, could then be demonstrated in a striking manner. And with a little trouble, specimens can be secured in which perfect regularity of response is found. The record given in fig. 16, obtained with a specimen of radish, shows how possible it is to secure plants in which response is absolutely regular. I subjected this to uniform stimulation at intervals of one minute, during half an hour, without detecting the least variation in the responses. But it is of course easier to find others in which the responses as a whole may be taken as regular, though there may be slight rhythmic fluctuations. And even in these cases the effect of reagents is too marked and sudden to escape notice.
The most important test for distinguishing vital phenomena is how they respond to narcotics and poisons. For instance, a nerve that has been numbed by chloroform shows a reduced response as the anesthetic takes effect. (See below, fig. 43.) Likewise, various poisons can permanently eliminate all responses. Strong alkalis and strong acids can kill a nerve. I have already demonstrated that plants that previously responded strongly do not respond at all after being treated with an anesthetic or poison. In these cases, only the final stage could be observed. However, it seemed important to trace the gradual effects of anesthesia or poisoning throughout the process. There were, however, two challenges that initially seemed difficult to overcome. First, we needed to find a specimen that normally showed no fatigue and could produce a consistent series of responses for a long time. The immediate changes in response due to the application of chemical reagents could then be clearly demonstrated. With a bit of effort, we can find specimens that exhibit perfect regularity in response. The data recorded in fig. 16, collected from a radish specimen, shows how possible it is to find plants with absolutely regular responses. I subjected this specimen to consistent stimulation every minute for half an hour without noticing any variation in the responses. However, it is generally easier to find others where the overall responses can be considered regular, even if there are slight rhythmic fluctuations. In these cases, the effects of reagents are too pronounced and sudden to go unnoticed.

Fig. 43.—Effect of Chloroform on Nerve Response (Waller)
Fig. 43.—Effect of Chloroform on Nerve Response (Waller)
For the obtaining of constant and strong response I found the best materials to be carrot and radish, selected individuals from which gave most satisfactory results. The carrots were at their best in August and September, after which their sensitiveness rapidly declined. Later, being obliged to seek for other specimens, I came upon radish, which gave good results in the early part of November; but the setting-in of the frost had a prejudicial effect on its responsiveness. Less perfect than these, but still serviceable, are the leaf-stalks of turnip and cauliflower. In these the successive responses as a whole may be regarded as regular, though a curious alternation is sometimes noticed, which, however, has a regularity of its own.
To get a consistent and strong response, I found the best materials to be carrots and radishes, with select individual specimens yielding the most satisfactory results. The carrots were at their peak in August and September, after which their sensitivity quickly declined. Later, needing to find other options, I came across radishes, which produced good results in early November; however, the onset of frost negatively affected their responsiveness. Less ideal but still useful are the leaf stalks of turnip and cauliflower. In these, the overall responses can be considered regular, although a curious alternation is sometimes observed, which still follows its own pattern.
My second misgiving was as to whether the action of reagents would be sufficiently rapid to display itself within the time limit of a photographic record. This would of course depend in turn upon the rapidity with which the tissues of the plant could absorb the reagent and be affected by it. It was a surprise to me to find that, with good specimens, the effect was manifested in the course of so short a time as a minute or so.
My second concern was whether the reagents would work fast enough to show results in the time it takes to capture a photographic record. This, of course, would depend on how quickly the plant's tissues could absorb the reagent and react to it. I was surprised to find that, with good specimens, the effect appeared in as little as a minute.
Effect of chloroform.—In studying the effect of chemical reagents in plants, the method is precisely similar to that employed with nerve; that is to say, where vapour of chloroform is used, it is blown into the plant chamber. In cases of liquid reagents, they are applied on the points of contact A and B and their close neighbourhood. The mode of experiment was (1) to obtain a series of normal responses to uniform stimuli, applied at regular intervals of time, say one minute, the record being taken the while on a photographic plate. (2) Without interrupting this procedure, the anæsthetic agent, vapour of chloroform, was blown into the closed chamber containing the plant. It will be seen how rapidly chloroform produces depression of response (fig. 44), and how the effect grows with time. In these experiments with plants, the same curious shifting of the zero line is sometimes noticed as in nerve when subjected similarly to the action of reagents. This is a point of minor importance, the essential point to be noticed being that the responses are rapidly reduced.
Effect of chloroform.—When studying how chemical substances affect plants, the method is quite similar to what’s used for nerves. In this case, the vapor of chloroform is introduced into the plant chamber. For liquid substances, they are applied to contact points A and B and the surrounding area. The experimental procedure was (1) to measure a series of normal responses to consistent stimuli applied at regular intervals, like one minute, with the results being recorded on a photographic plate. (2) Without stopping this process, the anesthetic vapor of chloroform was introduced into the closed chamber containing the plant. It will be evident how quickly chloroform causes a decrease in response (fig. 44), and how this effect intensifies over time. In these plant experiments, similar unexpected shifts of the zero line are sometimes observed as in nerve experiments when subjected to the action of reagents. This is a minor detail; the key takeaway is that the responses decrease rapidly.

Fig. 44.—Effect of Chloroform on Responses of Carrot
Fig. 44.—Impact of Chloroform on Carrot Responses
Stimuli of 25° vibration at intervals of one minute.
Stimuli of 25° vibration every minute.
Effects of chloral and formalin.—I give below (figs. 45, 46) two sets of records, one for the reagent chloral and the other for formalin. The reagents were applied in the form of a solution on the tissue at the two leading contacts, and the contiguous surface. The rhythmic fluctuation in the normal response shown in fig. 45 is interesting. The abrupt decline, within a minute of the application of chloral, is also extremely well marked.
Effects of chloral and formalin.—I am providing below (figs. 45, 46) two sets of records, one for the reagent chloral and the other for formalin. The reagents were applied as a solution on the tissue at the two main contact points and the surrounding surface. The rhythmic changes in the normal response shown in fig. 45 are interesting. The sharp drop, within a minute of applying chloral, is also very clearly observed.

Fig. 45.—Action of Chloral Hydrate on the Responses of Leaf-stalk of Cauliflower
Fig. 45.—Effect of Chloral Hydrate on the Responses of Cauliflower Leaf Stalk
Vibration of 25° at intervals of one minute.
Vibration of 25° per minute.

Response unaffected by variation of resistance.—In order to bring out clearly the main phenomena, I have postponed till now the consideration of a point of some difficulty. To determine the influence of a reagent in modifying the excitability of the tissue, we rely upon its effect in exalting or depressing the responsive E.M. variation. We read this effect by means of galvanometric deflections. And if the resistance of the circuit remained constant, then an increase of galvanometer deflection would accurately indicate a heightened or depressed E.M. response, due to greater or less excitability of tissue caused by the reagent. But, by the introduction of the chemical reagent, the resistance of the tissue may undergo change, and owing to this cause, modification of response as read by the galvanometer may be produced without any E.M. variation. The observed variation of response may thus be partly owing to some unknown change of resistance, as well as to that of the E.M. variation in response to stimulus.
Response unaffected by variation of resistance.—To clearly highlight the main phenomena, I have postponed discussing a point that can be a bit tricky. To determine how a reagent affects the excitability of the tissue, we look at its impact on increasing or decreasing the responsive E.M. variation. We measure this effect using galvanometric deflections. If the circuit's resistance stays constant, an increase in galvanometer deflection would accurately show a heightened or lowered E.M. response, resulting from the tissue's greater or lesser excitability due to the reagent. However, when the chemical reagent is introduced, the tissue's resistance can change, and this change can cause a modification in response as measured by the galvanometer, even without any E.M. variation. Therefore, the observed change in response may be partly due to an unknown change in resistance, in addition to the E.M. variation’s response to the stimulus.
We may however discriminate as to how much of the observed change is due to variation of resistance by comparing the deflections produced in the galvanometer by the action of a definite small E.M.F. before and after the introduction of the reagent. If the deflections be the same in both cases, we know that the resistance has not varied. If there have been any change, the variation of deflection will show the amount, and we can make allowance accordingly.
We can, however, determine how much of the observed change is due to variations in resistance by comparing the deflections produced in the galvanometer by a specific small E.M.F. before and after adding the reagent. If the deflections are the same in both cases, we know that the resistance hasn't changed. If there is any change, the difference in deflection will indicate the amount, and we can adjust for that.
I have however adopted another method, by which all necessity of correction is obviated, and the galvanometric deflections simply give E.M. variations, unaffected by any change in the resistance of the tissue. This is done by interposing a very large and constant resistance in the external circuit and thereby making other resistances negligible. An example will make this point clear. Taking a carrot as the vegetable tissue, I found its resistance plus the resistance of the non- polarisable electrode equal to 20,000 ohms. The introduction of a chemical reagent reduced it to 19,000 ohms. The resistance of the galvanometer is equal to 1,000 ohms. The high external resistance was 1,000,000 ohms. The variation of resistance produced in the circuit would therefore be 1,000 in (1,000,000 + 19,000 + 1,000) or one part in 1,020. Therefore the variation of galvanometric deflection due to change of resistance would be less than one part in a thousand (cf. fig. 49).
I have, however, taken a different approach that eliminates the need for correction, allowing the galvanometric deflections to represent E.M. variations without being affected by any changes in the tissue's resistance. This is accomplished by placing a very large and constant resistance in the external circuit, making other resistances insignificant. An example will clarify this point. Using a carrot as the vegetable tissue, I found its resistance, combined with the resistance of the non-polarizable electrode, to be 20,000 ohms. Adding a chemical reagent reduced it to 19,000 ohms. The resistance of the galvanometer is 1,000 ohms. The high external resistance was 1,000,000 ohms. The change in resistance within the circuit would therefore be 1,000 in (1,000,000 + 19,000 + 1,000) or one part in 1,020. Consequently, the change in galvanometric deflection due to the resistance change would be less than one part in a thousand (cf. fig. 49).
The advantage of the block method.—In these investigations I have used the block method, instead of that of negative variation, and I may here draw attention to the advantages which it offers. In the method of negative variation, one contact being injured, the chemical reagents act on injured and uninjured unequally, and it is conceivable that by this unequal action the resting difference of potential may be altered. But the intensity of response in the method of injury depends on this resting difference. It is thus hypothetically possible that on the method of negative variation there might be changes in the responses caused by variation of the resting difference, and not necessarily due to the stimulating or depressing effect of the reagent on the tissue.
The advantage of the block method.—In these investigations, I've used the block method instead of the negative variation method, and I want to highlight the benefits it provides. In the negative variation method, when one contact is damaged, the chemical reagents act differently on the injured and uninjured areas, which could change the resting potential difference. However, the intensity of response in the injury method relies on this resting difference. Therefore, it’s theoretically possible that in the negative variation method, the changes in responses could be due to variations in the resting difference, rather than the direct stimulating or depressing effect of the reagent on the tissue.
But by the block method the two contacts are made with uninjured surfaces, and the effect of reagents on both is similar. Thus no advantage is given to one contact over the other. The changes now detected in response are therefore due to no adventitious circumstance, but to the reagent itself. If further verification be desired as to the effect of the reagent, we can obtain it by alternate stimulation of the A and B ends. Both ends will then show the given change. I give below a record of responses given by two ends of leaf-stalk of turnip, stimulated alternately in the manner described. The stalk used was slightly conical, and owing to this difference between the A and B ends the responses given by one end were slightly different from those given by the other, though the stimuli were equal. A few drops of 10 per cent. solution of NaOH was applied to both the ends. It will be seen how quickly this reagent abolished the response of both ends (fig. 47).
But using the block method, the two contacts are made with intact surfaces, and the effect of the reagents on both is the same. So, there’s no advantage given to one contact over the other. The changes we see in response are therefore due to the reagent itself, not any outside factors. If we want to further confirm the effect of the reagent, we can do this by alternately stimulating the A and B ends. Both ends will then show the observed change. Below is a record of the responses from the two ends of a turnip leaf-stalk, stimulated alternately as described. The stalk used was slightly conical, and because of this difference between the A and B ends, the responses from one end were slightly different from the other, even though the stimuli were equal. A few drops of a 10 percent solution of NaOH were applied to both ends. It will be clear how quickly this reagent eliminated the response from both ends (fig. 47).

Fig. 47.—Abolition of Response at both A and B Ends by the Action of NaOH
Fig. 47.—Elimination of Response at both A and B Ends due to the Action of NaOH
Stimuli of 30° vibration were applied at intervals of one minute to A and B alternately. Response was completely abolished twenty-four minutes after application of NaOH.
Stimuli of 30° vibration were applied every minute to A and B alternately. The response was completely eliminated twenty-four minutes after applying NaOH.
Effect of dose.—It is sometimes found that while a reagent acts as a poison when given in large quantities, it may act as a stimulant in small doses. Of the two following records fig. 48 shows the slight stimulating effect of very dilute KOH, and fig. 49 exhibits nearly complete abolition of response by the action of the same reagent when given in stronger doses.
Effect of dose.—Sometimes a reagent that acts as a poison in large amounts can actually serve as a stimulant in smaller doses. Among the two records, fig. 48 demonstrates the mild stimulating effect of very dilute KOH, while fig. 49 shows that the same reagent completely blocks the response when administered in higher doses.

So we see that, judged by the final criterion of the effect produced by anæsthetics and poisons, the plant response fulfils the test of vital phenomenon. In previous chapters we have found that in the matter of response by negative variation, of the presence or absence of fatigue, of the relation between stimulus and response, of modification of response by high and low temperatures, and even in the matter of occasional abnormal variations such as positive response in a modified tissue, they were strictly correspondent to similar phenomena in animal tissues. The remaining test, of the influence of chemical reagents, having now been applied, a complete parallelism may be held to have been established between plant response on the one hand, and that of animal tissue on the other.
So we see that, based on the final criterion of the effects produced by anesthetics and poisons, the plant response meets the standards of a vital phenomenon. In previous chapters, we have found that regarding response by negative variation, the presence or absence of fatigue, the relationship between stimulus and response, the alteration of responses due to high and low temperatures, and even occasional abnormal variations like positive responses in modified tissues, these were all closely aligned with similar phenomena in animal tissues. Now that we have applied the last test, which examines the influence of chemical reagents, we can say that there is a complete parallel between plant responses and those of animal tissues.

Fig. 49.—Nearly Complete Abolition of Response by Strong KOH
Fig. 49.—Almost Complete Elimination of Response by Strong KOH
The two vertical lines are galvanometer deflections due to ·1 volt, before and after the application of reagent. It will be noticed that the total resistance remains unchanged.
The two vertical lines show the galvanometer readings due to 1 volt, before and after the reagent is applied. You'll notice that the total resistance stays the same.
CHAPTER X
RESPONSE IN METALS
- Is response found in inorganic substances?
- —Experiment on tin, block method
- —Anomalies of existing terminology
- —Response by method of depression
- —Response by method of exaltation.
We have now seen that the electrical sign of life is not confined to animals, but is also found in plants. And we have seen how electrical response serves as an index to the vital activity of the plant, how with the arrest of this vital activity electrical response is also arrested temporarily, as in the case amongst others of anæsthetic action, and permanently, for instance under the action of poisons. Thus living tissues—both animal and vegetable—may pass from a responsive to an irresponsive condition, from which latter there may or may not be subsequent revival.
We have now observed that the electrical signs of life are not limited to animals but can also be found in plants. We've also seen how electrical responses indicate the vital activities of a plant, and how, when these vital activities stop, the electrical responses temporarily stop as well, such as in the case of anesthetics, and permanently when affected by poisons. Thus, living tissues—both animal and plant—can shift from a responsive state to an unresponsive state, from which revival may or may not occur later.
Hitherto, as already said, electrical response in animals has been regarded as a purely physiological phenomenon. We have proved by various tests that response in plants is of the same character. And we have seen that by physiological phenomena are generally understood those of which no physical explanation can be offered, they being supposed to be due to the play of some unknown vital force existing in living substances and giving rise to electric response to stimulation as one of its manifestations.
So far, as mentioned before, electrical response in animals has been seen as strictly a physiological phenomenon. We've shown through various tests that the response in plants is the same. Generally, physiological phenomena are understood to be those for which no physical explanation can be provided; they are thought to be caused by some unknown vital force present in living things, resulting in electric responses to stimulation as one of its effects.
Is response found in inorganic substances?[14]—It is now for us, however, to examine into the alleged super-physical character of these phenomena by stimulating inorganic substances and discovering whether they do or do not give rise to the same electrical mode of response which was supposed to be the special characteristic of living substances. We shall use the same apparatus and the same mode of stimulation as those employed in obtaining plant response, merely substituting, for the stalk of a plant, a metallic wire, say ‘tin’ (fig. 50). Any other metal could be used instead of tin.
Is response found in inorganic substances?[14]—Now, we need to investigate the supposed super-physical nature of these phenomena by testing inorganic substances to see if they produce the same electrical response that was thought to be unique to living things. We'll use the same equipment and method of stimulation as we did to measure plant responses, only replacing the plant stalk with a metal wire, like ‘tin’ (fig. 50). Any metal could be used in place of tin.
Experiment on tin, block method.—Let us then take a piece of tin wire[15] from which all strains have been previously removed by annealing, and hold it clamped in the middle at C. If the strains have been successfully removed A and B will be found iso-electric, and no current will pass through the galvanometer. If A and B are not exactly similar, there will be a slight current. But this will not materially affect the results to be described presently, the slight existing current merely adding itself algebraically to the current of response.
Experiment on tin, block method.—Let's take a piece of tin wire[15] that has had all strains removed by annealing, and hold it clamped in the middle at C. If the strains have been successfully removed, A and B will be found to be iso-electric, and no current will pass through the galvanometer. If A and B are not exactly the same, there will be a slight current. However, this won't significantly impact the results that will be described shortly, as the minor existing current will simply add algebraically to the response current.
If we now stimulate the end A by taps, or better still by torsional vibration, a transitory ‘current of action’ will be found to flow in the wire from B to A, from the unstimulated to the stimulated, and in the galvanometer from the stimulated to the unstimulated. Stimulation of B will give rise to a current in an opposite direction.
If we now stimulate the end A by tapping it, or better still by twisting vibrations, a temporary ‘current of action’ will be found to flow in the wire from B to A, from the unstimulated end to the stimulated end, and in the galvanometer from the stimulated end to the unstimulated end. Stimulating B will create a current in the opposite direction.

Fig. 50.—Electric Response in Metals
Fig. 50.—Electrical Response in Metals
(a) Method of block; (b) Equal and opposite responses when the ends A and B are stimulated; the dotted portions of the curves show recovery; (c) Balancing effect when both the ends are stimulated simultaneously.
(a) Block method; (b) Equal and opposite responses when ends A and B are stimulated; the dotted sections of the curves indicate recovery; (c) Balancing effect when both ends are stimulated at the same time.
Experiment to exhibit the balancing effect.—If the wire has been carefully annealed, the molecular condition of its different portions is found to be approximately the same. If such a wire be held at the ‘balancing point’ (which is at or near the middle) by the clamp, and a quick vibration, say, of 90° be given to A, an upward deflection will be produced; if a vibration of 90° be given to B, there will be an equal downward deflection. If now both the ends A and B are vibrated simultaneously, the responsive E.M. variation at the two ends will continuously balance each other and the galvanometer spot will remain quiescent (fig. 30, A, B, R). This balance will be still maintained when the block is removed and the wire is vibrated as a whole. It is to be remembered that with the length of wire constant, the intensity of stimulus increases with the amplitude of vibration. Again, keeping the amplitude constant, the intensity of stimulus is increased by shortening the wire. Hence it will be seen that if the clamp be shifted from the balancing point towards A, simultaneous vibration of A and B through 90° will now give a resultant upward deflection, showing that the A response is now relatively stronger. Thus keeping the rest of the circuit untouched, merely moving the clamp from the left, past the balancing point to the right, we get either a positive, or zero, or negative, resultant effect.
Experiment to show the balancing effect.—If the wire has been carefully softened, the molecular structure of its different sections is found to be roughly the same. If you hold this wire at the 'balancing point' (which is around the middle) with the clamp, and give a quick vibration, say, of 90° to A, it will create an upward deflection; if you give a 90° vibration to B, there will be an equal downward deflection. If both ends A and B are vibrated at the same time, the responsive E.M. variation at both ends will continuously balance each other, and the galvanometer spot will stay steady (fig. 30, A, B, R). This balance will still hold when the block is removed and the wire is vibrated as a whole. It should be noted that with a constant length of wire, the intensity of the stimulus increases with the amplitude of vibration. Additionally, keeping the amplitude constant, the intensity of the stimulus is increased by shortening the wire. Therefore, if the clamp is moved from the balancing point towards A, simultaneous vibration of A and B through 90° will now produce a net upward deflection, indicating that the A response is now relatively stronger. Thus, with the rest of the circuit unchanged, simply moving the clamp from the left, past the balancing point to the right, we can observe either a positive, zero, or negative resultant effect.
In tin the current of response is from the less to the more excited point. In the retina also, we found the current of action flowing from the less stimulated to the more stimulated, and as that is known as a positive response, we shall consider the normal response of tin to be in like manner positive.
In the current of response, it goes from the less excited point to the more excited one. In the retina too, we observed the action flowing from the less stimulated to the more stimulated areas, and since that is known as a positive response, we will consider the normal response of tin to also be a positive one.
Just as the response of retina or nerve, under certain molecular conditions, undergoes reversal, the positive being then converted into negative, and negative into positive, so it will be shown that the response in metallic wires under certain conditions is found to undergo reversal.
Just as the response of the retina or nerve, under certain molecular conditions, can reverse—turning positive into negative and negative into positive—it's been demonstrated that the response in metallic wires under certain conditions also experiences reversal.
Anomalies of present terminology.—When there is no current of injury, a particular current of response can hardly be called a negative, or positive, variation. Such nomenclature is purely arbitrary, and leads, as will be shown, to much confusion. A more definite terminology, free from misunderstanding, would be, as already said, to regard the current towards the more stimulated as positive, and that towards the less stimulated, in tissue or wire, as negative.
Issues with Current Terminology.—When there isn't a consistent flow of injury, it’s difficult to classify a specific flow of response as either negative or positive variation. This classification is completely arbitrary and, as will be shown, leads to a lot of confusion. A clearer way to think about it, as mentioned earlier, would be to consider the current moving towards the more stimulated area as positive, and the current moving towards the less stimulated area, whether in tissue or wire, as negative.
The stimulated end of tin, say the end A, thus becomes zincoid, i.e. the current through the electrolyte (non-polarisable electrodes with interposed galvanometer) is from A to B, and through the wire, from the less stimulated B to the more stimulated A. Conversely, when B is stimulated, the action current flows round the circuit in an opposite direction. This positive is the most usual form of response, but there are cases where the response is negative.
The activated end of tin, let’s call it end A, then becomes zincoid, meaning the current through the electrolyte (with non-polarizable electrodes and an integrated galvanometer) flows from A to B, and through the wire, from the less activated B to the more activated A. On the other hand, when B is activated, the action current travels around the circuit in the opposite direction. This positive response is the most common, but there are cases where the response is negative.
In order to show that normally speaking a stimulated wire becomes zincoid, and also to show once more the anomalies into which we may fall by adopting no more definite terminology than that of negative variation, I have devised the following experiment (fig. 51). Let us take a bar, one half of which is zinc and the other half copper, clamped in the middle, so that a disturbance produced at one end may not reach the other; the two ends are connected to a galvanometer through non-polarisable electrodes. The current through the electrolyte (non-polarisable electrodes and interposed galvanometer) will then flow from left to right. We must remember that metals under stimulation generally become, in an electrical sense, more zinc-like. On vibrating the copper end (inasmuch as copper would then become more zinc-like) the difference of potential between zinc and copper ought to be diminished, and the current flowing in the circuit would therefore be lessened. But vibration of the zinc end ought to increase the potential difference, and there ought to be then an increase of current during stimulation of zinc.
To show that under normal conditions a stimulated wire acts more like zinc, and to highlight again the problems we may face by using vague terminology like negative variation, I’ve set up the following experiment (__A_TAG_PLACEHOLDER_0__). Let’s take a bar that’s zinc on one half and copper on the other, clamped in the middle so that a disturbance at one end won’t affect the other. The two ends are connected to a galvanometer using non-polarizable electrodes. The current through the electrolyte (with non-polarizable electrodes and the galvanometer in between) will then flow from left to right. We should keep in mind that metals usually become more zinc-like when stimulated. When we vibrate the copper end (since copper becomes more zinc-like), the potential difference between zinc and copper should decrease, leading to a reduced current in the circuit. However, vibrating the zinc end should increase the potential difference, resulting in a higher current during the stimulation of zinc.

Fig. 51.—Current of Response towards the Stimulated End
Fig. 51.—Response Flow toward the Stimulated End
Hence when Cu stimulated: action current →, normal E.M.F. diminished (·85-·009) V.
Hence when Cu stimulated: action current →, normal E.M.F. diminished (·85-·009) V.
When Zn stimulated: action current ←, normal E.M.F. increased (·85 + ·013) V.
When Zn was stimulated: action current ←, normal E.M.F. increased (·85 + ·013) V.
In the particular experiment of fig. 51, the E.M.F. between the zinc and copper ends was found to be ·85 volt. This was balanced by a potentiometer arrangement, so that the galvanometer spot came to zero. On vibrating the zinc wire, a deflection of 33 dns. was obtained, in a direction which showed an increase of E.M.F. On stopping the vibration, the spot of light came back to zero. On now vibrating the copper wire, a deflection of 23 dns. was obtained in an opposite direction, showing a diminution of E.M.F. This transitory responsive variation disappeared on the cessation of disturbance.
In the experiment of __A_TAG_PLACEHOLDER_0__, the electromotive force (E.M.F.) between the zinc and copper ends was recorded at 0.85 volts. This was balanced using a potentiometer setup, which brought the galvanometer reading to zero. When the zinc wire was vibrated, a deflection of 33 dns. was noted, indicating an increase in E.M.F. Once the vibration stopped, the light spot returned to zero. When the copper wire was vibrated, a deflection of 23 dns. was observed in the opposite direction, indicating a diminution in E.M.F. This temporary change disappeared when the vibration ceased.
By disturbing the balance of the potentiometer, the galvanometer deflection due to a known increase of E.M.F. was found from which the absolute E.M. variation caused by disturbance of copper or zinc was determined.
By disrupting the balance of the potentiometer, we measured the galvanometer's deflection due to a known increase in E.M.F., which allowed us to determine the exact E.M.F. variation caused by the disturbance of copper or zinc.
It was thus found that stimulation of zinc had increased the P.D. by fifteen parts in 1,000, whereas stimulation of copper had decreased it by eleven parts in 1,000. According to the old terminology, the response due to stimulation of zinc would have been regarded as positive variation, that of copper negative. The responses however are not essentially opposite in character, the action current in the bar being in both cases towards the more excited. For this reason it would be preferable, as already said, to employ the terms positive and negative in the sense I have suggested, i.e. positive, when the current in the acted substance is towards the more excited, and negative, when towards the less excited. The method of block is, as I have already shown, the most perfect for the study of these responses.
It was found that vibrating zinc increased the potential difference (P.D.) by fifteen parts in 1,000, while vibrating copper decreased it by eleven parts in 1,000. In older terminology, the response from zinc stimulation would have been seen as a positive variation, and that from copper as negative. However, these responses are not fundamentally opposite; in both cases, the action current in the material is directed towards the more excited state. For this reason, as mentioned earlier, it would be better to use the terms positive and negative as I suggested: positive when the current in the substance being affected is toward the more excited state, and negative when it is toward the less excited state. The block method, as I have already shown, is the most effective way to study these responses.
In the experiment fig. 50, if the block is abolished and the wire is struck in the middle, a wave of molecular disturbance will reach A and B. The mechanical and the attendant electrical disturbance will at these points reach a maximum and then gradually subside. The resultant effect in the galvanometer will be due to EA-EB when EA and EB are the electrical variations produced at A and B by the stimulus. The electric changes at A and B will continuously balance each other, and the resultant effect on the galvanometer will be zero: (a) if the exciting disturbance reaches A and B at the same time and with the same intensity; (b) if the molecular condition is similar at the two points; and (c) if the rate of rise and subsidence of excitation is the same at the two points. In order that a resultant effect may be exhibited in the galvanometer, matters have to be so arranged that the disturbance may reach one point, say A, and not B, and vice versa. This was accomplished by means of a clamp, in the method of block. Again a resultant differential action may be obtained even when the disturbance reaches both A and B, if the electrical excitability of one point is exalted or depressed by physical or chemical means. We shall in Chap. XVI study in detail the effect of chemical reagents in producing the enhancement or depression of excitability. There are thus two other means of obtaining a resultant effect—(2) by the method of relative depression, (3) by the method of relative exaltation.
In the experiment fig. 50, if the block is removed and you hit the wire in the middle, a wave of molecular disturbance will reach A and B. The mechanical and related electrical disturbance will peak at these points and then gradually decrease. The overall effect in the galvanometer will stem from EA-EB, with EA and EB representing the electrical changes produced at A and B by the stimulus. The electrical changes at A and B will continuously balance each other out, resulting in a zero effect on the galvanometer: (a) if the exciting disturbance hits A and B at the same time and with equal intensity; (b) if the molecular conditions are similar at both points; and (c) if the rate of increase and decrease of excitation is the same at both points. For a resultant effect to be displayed on the galvanometer, it must be set up so that the disturbance reaches one point, like A, but not B, and vice versa. This was achieved using a clamp in the blocking method. Additionally, a resultant differential action can occur even when the disturbance reaches both A and B, if the electrical excitability at one point is raised or lowered by physical or chemical means. We will explore the effects of chemical reagents on enhancing or suppressing excitability in detail in Chap. XVI. Thus, there are two more ways to achieve a resultant effect: (2) through the method of relative depression, and (3) through the method of relative exaltation.
Electric response by method of depression.—We may thus by reducing or abolishing the excitability of one end by means of suitable chemical reagents (so-called method of injury) obtain response in metals without a block. The entire length of the wire may then be stimulated and a resultant response will be produced, owing to the difference between the excitability of the two ends. A piece of tin wire is taken, and one normal contact is made at A (strip of cloth moistened with water, or very dilute salt solution). The excitability of B is depressed by a few drops of strong potash or oxalic acid. By the application of the latter there will be a small P.D. between A and B; this will simply produce a displacement of zero. By means of a potentiometer the galvanometer spot may be brought back to the original position. The shifting of the zero will not affect the general result. The effect of mechanical stimulus is to produce a transient electro-motive response, which will be superposed algebraically on the existing P.D. The deflection will take place from the modified zero to which the spot returns during recovery. On now stimulating the wire as a whole by, say, torsional vibration, the current of response will be found towards the more excitable, i.e. from B to A (fig. 52, a).
Electric response through depression method.—By reducing or eliminating the excitability of one end using appropriate chemical agents (known as the injury method), we can get a response in metals without obstruction. The entire length of the wire can then be stimulated, and a resulting response will occur due to the difference in excitability between the two ends. We take a piece of tin wire and establish a normal contact at A (using a strip of cloth moistened with water or a very dilute salt solution). The excitability of B is lowered by applying a few drops of strong potash or oxalic acid. With the latter applied, there will be a small potential difference (P.D.) between A and B; this will simply create a displacement of zero. A potentiometer can be used to return the galvanometer spot to the original position. This shift from zero will not impact the overall result. The mechanical stimulus will produce a temporary electro-motive response, which will algebraically add to the existing P.D. The deflection will occur from the modified zero point back to which the spot returns during recovery. Now, when stimulating the wire as a whole, for example, with torsional vibration, the response current will flow toward the more excitable end, meaning from B to A (fig. 52, a).
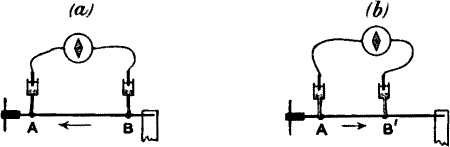
Fig. 52.—Response by Method of Depression (Without Block)
Fig. 52.—Response by Method of Depression (Without Block)
When the wire is stimulated as a whole the current of response is towards the more excitable.
When the wire is stimulated as a whole, the current response moves toward the more excitable part.
In (a) A is a normal contact, B has been depressed by oxalic acid; current of response is towards the more excitable A.
In (a) A is a normal contact, B has been depressed by oxalic acid; the response moves towards the more excitable A.
In (b) the same wire is used, only A is depressed by oxalic acid and a normal contact is made at a fresh point B′, a little to the left of B in (a). Current of response is now from A towards the more excitable B′.
In (b), the same wire is used, but A is pressed down by oxalic acid and a normal contact is made at a new point B′, slightly to the left of B in (a). The current of response is now from A to the more excitable B′.
A corroborative reversal experiment may next be made on the same piece of wire. The normal contact, through water or salt solution, is now made at B′, a little to the left of B. The excitability of A is now depressed by oxalic acid. On stimulation of the whole wire, the current of response will now be found to flow in an opposite direction—i.e. from A to B′—but still from the relatively less to the relatively more excitable (fig. 52, b).
A confirming reversal experiment can be conducted on the same piece of wire next. The standard contact, using water or a salt solution, is now made at B', slightly to the left of B. The excitability of A is now reduced by oxalic acid. When the entire wire is stimulated, the response current will now be found flowing in the opposite direction—i.e., from A to B′—but still from the relatively less excitable to the relatively more excitable (fig. 52, b).

Fig. 53.—Method of Exaltation
Fig. 53.—Exaltation Method
The contact B is made more excitable by chemical stimulant (Na2CO3). The current of response is towards the more excitable B.
The contact B is made more excitable by the chemical stimulant (Na2CO3). The response current is directed toward the more excitable B.
Method of exaltation.—A still more striking corroboration of these results may, however, be obtained by the converse process of relative exaltation of the responsiveness of one contact. This may be accomplished by touching one contact, say B, with a reagent which like Na2CO3 exalts the electric excitability. On stimulation of the wire, the current of response is towards the more excitable B (fig. 53).
Method of Exaltation.—An even more striking confirmation of these results can be obtained by the opposite method of enhancing the responsiveness of one contact. This can be done by applying a reagent, such as Na2CO3, to one contact, let’s say B, which boosts its electric excitability. When the wire is stimulated, the response current will flow towards the more excitable B (fig. 53).
I give four records (fig. 54) which will clearly exhibit the responses as obtained by the methods of relative depression or exaltation. In (a) B is touched with the excitant Na2CO3, a permanent current flows from A to B, response to stimulus is in the same direction as the permanent current (positive variation). In (b) B is touched with a trace of the depressant oxalic acid, the permanent current is in the same direction as before, but the current of response is in the opposite direction (negative variation). In (c) B is touched with dilute KHO, the response is exhibited by a positive variation. In (d) B is touched with strong KHO, the response is now exhibited by a negative variation. The last two results, apparently anomalous, are due to the fact, which will be demonstrated later, that KHO in minute quantities is an excitant, while in large quantities it is a depressant.
I provide four records (fig. 54) that will clearly show the responses obtained through the methods of relative depression or exaltation. In (a), B is treated with the excitant Na2CO3, resulting in a permanent current flowing from A to B, and the response to the stimulus aligns with the direction of the permanent current (positive variation). In (b), B is treated with a trace of the depressant oxalic acid; the permanent current still flows in the same direction as before, but the response current flows in the opposite direction (negative variation). In (c), B is treated with dilute KHO, and the response shows a positive variation. In (d), B is treated with strong KHO, and now the response shows a negative variation. The last two results, which seem unusual, are due to the fact that KHO in small amounts acts as an excitant, while in larger amounts it acts as a depressant, a fact that will be demonstrated later.

Fig. 54
Fig. 54
Current of response is always towards the more excitable point. The current of response always moves towards the more exciting point. |
The response is up when B is more excitable, and down when A is more excitable. The response goes up when B is more excited and goes down when A is more excited. Lines thus ------ indicate deflection due to permanent current. Lines thus ------ indicate deflection caused by constant current. |
We have thus seen that we may obtain response (1) by block method, (2) by the method of injury, or relative depression of responsiveness of one contact, and (3) by the method of relative exaltation of responsiveness of one contact. In all these cases alike we obtain a consistent action current, which in tin is normally positive, or towards the relatively more excited.
We have seen that we can achieve response (1) using the block method, (2) by applying the method of injury or by lowering the responsiveness of one contact, and (3) through the method of increasing the responsiveness of one contact. In all these cases, we produce a consistent action current, which in tin is usually positive, or directed towards the relatively more excited.
FOOTNOTES:
[14] Following another line of inquiry I obtained response to electric stimulus in inorganic substances using the method of conductivity variation (see ‘De la Généralité des Phénomènes Moléculaires Produits par l’Electricité sur la Matière Inorganique et sur la Matière Vivante,’ Travaux du Congrès International de Physique, Paris, 1900; and also ‘On Similarities of Effect of Electric Stimulus on Inorganic and Living Substances,’ British Association 1900. See Electrician). To bring out the parallelism in all details between the inorganic and living response, I have in the following chapters used the method of electro-motive variation employed by physiologists.
[14] Following another line of inquiry, I received responses to electric stimuli in inorganic substances using the method of conductivity variation (see ‘De la Généralité des Phénomènes Moléculaires Produits par l’Electricité sur la Matière Inorganique et sur la Matière Vivante,’ Travaux du Congrès International de Physique, Paris, 1900; and also ‘On Similarities of Effect of Electric Stimulus on Inorganic and Living Substances,’ British Association 1900. See Electrician). To highlight the similarities in detail between the inorganic and living responses, I have used the method of electro-motive variation applied by physiologists in the following chapters.
CHAPTER XI
INORGANIC RESPONSE—MODIFIED APPARATUS TO EXHIBIT RESPONSE IN METALS
- Conditions of obtaining quantitative measurements
- —Modification of the block method
- —Vibration cell
- —Application of stimulus
- —Graduation of the intensity of stimulus
- —Considerations showing that electric response is due to molecular disturbance
- —Test experiment
- —Molecular voltaic cell.
We have already seen that metals respond to stimulus by E.M. variation, just as do animal and vegetable tissues. We have yet to see whether the similarity extends to this point only, or goes still further, whether the response-curves of living and in organic are alike, and whether the inorganic response-curve is modified, as living response was found to be, by the influence of external agencies. If so, are the modifications similar? What are the effects of superposition of stimuli? Is there fatigue? If there be, in what way does it affect the curves? And lastly, is the response of metals exalted or depressed by the action of chemical reagents?
We’ve already seen that metals react to stimuli through electromagnetic variation, just like animal and plant tissues do. Now we need to determine whether this similarity is limited to just that aspect or extends further. Are the response curves of living organisms and inorganic materials comparable, and is the inorganic response curve influenced, like the living response, by external factors? If that’s the case, are the changes similar? What happens with the overlapping of stimuli? Is there such a thing as fatigue? If it exists, how does it impact the curves? Lastly, do chemical reagents enhance or diminish the response of metals?
Conditions of obtaining quantitative measurements.—In order to carry out these investigations, it is necessary to remove all sources of uncertainty, and obtain quantitative measurements. Many difficulties at first presented themselves in the course of this attempt, but they were completely removed by the adoption of the following experimental modification. In the simple arrangement for qualitative demonstration of response in metals previously described, successive experiments will not give results which are strictly comparable (1) unless the resistance of the circuit be maintained constant. This would necessitate the adoption of some plan for keeping the electrolytic contacts at A and B absolutely invariable. There should then be no chance of any shifting or variation of contact. (2) There must also be some means of applying successive stimuli of equal intensity. (3) And for certain further experiments it will be necessary to have some way of gradually increasing or decreasing the stimuli in a definite manner.
Conditions for Obtaining Quantitative Measurements.—To conduct these investigations, it's essential to eliminate all sources of uncertainty and obtain quantitative measurements. Many challenges initially arose during this effort, but they were completely resolved by implementing the following experimental adjustments. In the simple setup for qualitatively demonstrating the response in metals described earlier, consecutive experiments won't yield results that are directly comparable (1) unless the circuit's resistance remains constant. This would require a method to keep the electrolytic contacts at A and B absolutely unchanged. There must be no possibility of any shifting or variation in contact. (2) Additionally, there needs to be a way to apply successive stimuli of equal intensity. (3) For certain additional experiments, it will also be necessary to have a means of gradually increasing or decreasing the stimuli in a controlled manner.
Modification of the block method.—By consideration of the following experimental modifications of the block method (fig. 55), it will be found easy to construct a perfected form of apparatus, in which all these conditions are fully met. The essentials to be kept in mind were the introduction of a complete block midway in the wire, so that the disturbance of one half should be prevented from reaching the other, and the making of a perfect electrolytic contact for the electrodes leading to the galvanometer.
Modification of the block method.—By looking at the following experimental changes to the block method (fig. 55), it will be easy to create an improved version of the equipment, in which all these requirements are fully met. The key points to remember were the addition of a complete block in the middle of the wire, so that the disturbance of one half wouldn't affect the other, and ensuring a perfect electrolytic connection for the electrodes going to the galvanometer.
Starting from the simple arrangement previously described where a straight wire is clamped in the middle (fig. 55, a), we next arrive at (b). Here the wire A B is placed in a U tube and clamped in the middle by a tightly fitting cork. Melted paraffin wax is poured to a certain depth in the bend of the tube. The two limbs of the tube are now filled with water, till the ends A and B are completely immersed. Connection is made with the non-polarisable electrodes by the side tubes. Vibration may be imparted to either A or B by means of ebonite clip holders seen at the upper ends A B of the wire.
Starting from the basic setup described earlier, where a straight wire is clamped in the middle (fig. 55, a), we move on to (b). In this setup, the wire A B is placed in a U tube and secured in the middle with a tightly fitting cork. Melted paraffin wax is poured to a specific depth in the bend of the tube. The two ends of the tube are filled with water until the ends A and B are fully submerged. Connection is established with the non-polarizable electrodes via the side tubes. Vibration can be applied to either A or B using ebonite clip holders located at the upper ends A B of the wire.

Fig. 55.—Successive Modifications of the Block Method from the ‘Straight Wire’ (a) to ‘Cell Form’ (e)
Fig. 55.—Successive Modifications of the Block Method from the ‘Straight Wire’ (a) to ‘Cell Form’ (e)
When A is excited, current of response in the wire is from less excited B to more excited A. Note that though the current of response is constant in direction, the galvanometer deflection in (d) will be opposite to that in (b).
When A is excited, the flow of response in the wire goes from the less excited B to the more excited A. It's important to note that even though the flow of response is consistent in direction, the galvanometer deflection in (d) will be opposite to that in (b).
It will be seen that the two limbs of the tube filled with water serve the purpose of the strip of moistened cloth used in the last experiment to make electric connections with the leading-out electrodes—with the advantage that we have here no chance of any shifting of contact or variation of surface, the contact between the wire and the surrounding liquid being perfect and invariable.
It can be observed that the two sides of the tube filled with water function like the damp cloth used in the previous experiment to connect with the output electrodes—offering the advantage that there’s no risk of any movement or changes in surface, as the connection between the wire and the surrounding liquid is consistent and stable.
On now vibrating the end A of the tin wire by means of the ebonite clip holder, a current will be found to flow from B to A through the wire—that is to say, towards the excited—and from A to B in the galvanometer.
On vibrating the end A of the tin wire using the ebonite clip holder, a current will flow from B to A through the wire—that is, towards the excited end—and from A to B in the galvanometer.
The next modification (c) is to transfer the galvanometer from the electrolytic to the metallic part of the circuit, that is to say, it is interposed in a gap made by cutting the wire A B, the upper part of the circuit being directly connected by the electrolyte. Vibration of A will now give rise to a current of response which flows in the metallic part of the circuit with the interposed galvanometer from B to A. We see that though the direction of the current in this is the same as in the last case, yet the galvanometer deflection is now reversed, for the evident reason that we have it interposed in the metallic and not in the electrolytic part of the circuit.
The next change (c) is to move the galvanometer from the electrolytic section to the metallic part of the circuit. This means it is placed in the gap created by cutting the wire A B, with the upper part of the circuit connected directly through the electrolyte. Vibration of A will now produce a response current that flows through the metallic part of the circuit with the galvanometer in between B and A. We can see that even though the direction of the current is the same as in the previous case, the deflection of the galvanometer is now reversed. This is clearly because it is placed in the metallic section instead of the electrolytic part of the circuit.
The next arrangement (d) consists simply of the preceding placed upside down. Here A and B are held parallel to each other in an electrolytic bath (water). Mechanical vibration may now be applied to A without affecting B, and vice versa.
The next setup (d) is just the previous one flipped upside down. Here, A and B are kept parallel to each other in an electrolytic bath (water). You can now apply mechanical vibrations to A without impacting B, and the same goes for the reverse.
The actual apparatus, of which this is a diagrammatic representation, is seen in (e).
The actual device, which is shown in this diagram, can be found in (e).
Two pieces, from the same specimen of wire, are clamped separately at their lower ends by means of ebonite screws, in an L-shaped piece of ebonite. The wires are fixed at their upper ends to two electrodes —leading to the galvanometer—and kept moderately and uniformly stretched by spiral springs. The handle, by which a torsional vibration is imparted to the wire, may be slipped over either electrode. The amplitude of vibration is measured by means of a graduated circle.
Two pieces of wire, taken from the same specimen, are separately clamped at their lower ends using ebonite screws in an L-shaped ebonite piece. The upper ends of the wires are attached to two electrodes —which connect to the galvanometer—and are kept moderately and uniformly stretched by spiral springs. The handle for creating a torsional vibration in the wire can slide over either electrode. The amplitude of the vibration is measured using a graduated circle.
It will be seen from these arrangements:
It will be clear from these arrangements:
(1) That the cell depicted in (e) is essentially the same as that in (a).
(1) The cell shown in (e) is basically the same as the one in (a).
(2) That the wires in the cell being immersed to a definite depth in the electrolyte there is always a perfect and invariable contact between the wire and the electrolyte. The difficulty as regards variation of contact is thus eliminated.
(2) The wires in the cell are submerged to a specific depth in the electrolyte, ensuring there is always a consistent and unchanging contact between the wire and the electrolyte. This eliminates any issues related to variations in contact.

Fig. 56.—Equal and Opposite Responses exhibited by A and B
Fig. 56.—Equal and Opposite Responses shown by A and B
(3) That as the wires A and B are clamped separately below, we may impart a sudden molecular disturbance to either A or B by giving a quick to-and-fro (torsional) vibration round the vertical wire, as axis, by means of the handle. As the wire A is separate from B, disturbance of one will not affect the other. Vibration of A produces a current in one direction, vibration of B in the opposite direction. Thus we have means of verifying every experiment by obtaining corroborative and reversed effects. When the two wires have been brought to exactly the same molecular condition by the processes of annealing or stretching, the effects obtained on subjecting A or B to any given stimulus are always equal (fig. 56).
(3) Since the wires A and B are secured separately below, we can create a sudden molecular disturbance in either A or B by giving a quick back-and-forth (torsional) vibration around the vertical wire, using the handle. Because wire A is separate from B, disturbing one won't impact the other. Vibrating A generates a current in one direction, while vibrating B generates a current in the opposite direction. This gives us a way to verify every experiment by obtaining consistent and opposing effects. When both wires have been brought to exactly the same molecular state through the processes of annealing or stretching, the results from applying any given stimulus to A or B are always the same (fig. 56).
Usually I interpose an external resistance varying from one to five megohms according to the sensitiveness of the wire. The resistance of the electrolyte in the cell is thus relatively small, and the galvanometer deflections are proportional to the E.M. variations. It is always advisable to have a high external resistance, as by this means one is not only able to keep the deflections within the scale, but one is not troubled by slight accidental disturbances.
Usually, I add an external resistance ranging from one to five megohms depending on how sensitive the wire is. This keeps the resistance of the electrolyte in the cell relatively low, and the galvanometer readings are proportional to the E.M. changes. It's always a good idea to use a high external resistance because this way, you can keep the readings within the scale, and you won't be bothered by minor accidental disturbances.
Graduation of intensity of stimulus.—If now a rapid torsional vibration be given to A or B, an E.M. variation will be induced. If the amplitude of vibration be kept constant, successive responses—in substances which, like tin, show no fatigue—will be found to be absolutely identical. But as ‘the amplitude of vibration’ is increased, response will also become enhanced (see Chap. XV).
Graduation of intensity of stimulus.—If a rapid twisting vibration is applied to A or B, it will induce an E.M. variation. If the vibration's amplitude stays the same, the responses in materials like tin, which don't show fatigue, will be exactly the same every time. However, as the 'amplitude of vibration' is increased, the response will also get stronger (see Chap. XV).
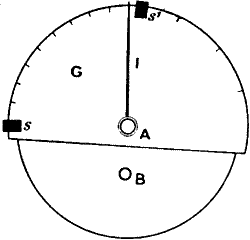
Fig. 57.—Top View of the Vibration Cell
Fig. 57.—Top View of the Vibration Cell
The amplitude of vibration is determined by means of movable stops S S′, fixed to the edge of the graduated circle G. The index arm I plays between the stops. (The second index arm, connected with B, and the second circle are not shown.)
The amplitude of vibration is measured using movable stops S S′, which are attached to the edge of the graduated circle G. The index arm I moves between the stops. (The second index arm, connected with B, and the second circle are not shown.)
Amplitude of vibration is measured by means of the graduated circle (fig. 57). A projecting index, in connection with the vibration-head, plays between fixed and sliding stops (S and S′), one at the zero point of the scale, and the other movable. The amplitude of a given vibration can thus be predetermined by the adjustment of the sliding stop. In this way we can obtain either uniform or definitely graduated stimuli.
The amplitude of vibration is measured using a graduated circle (fig. 57). A projecting index, connected to the vibration head, moves between fixed and sliding stops (S and S'), with one at the zero point of the scale and the other adjustable. The amplitude of a specific vibration can therefore be set in advance by adjusting the sliding stop. This allows us to create either uniform or precisely graduated stimuli.
Considerations showing that electric response is due to molecular disturbance.—The electromotive variation varies with the substance. With superposition of stimuli, a relatively high value is obtained in tin, amounting sometimes to nearly half a volt, whereas in silver the electromotive variation is only about ·01 of this value. The intensity of the response, however, does not depend on the chemical activity of the substance, for the electromotive variation in the relatively chemically inactive tin is greater than that of zinc. Again, the sign of response, positive or negative, is sometimes modified by the molecular condition of the wire (see Chap. XII).
Considerations showing that electric response is due to molecular disturbance.—The electromotive variation changes depending on the substance. When multiple stimuli are applied, a relatively high value is seen in tin, reaching nearly half a volt at times, whereas in silver, the electromotive variation is only about ·01 of that value. However, the intensity of the response does not rely on the chemical activity of the substance, since the electromotive variation in relatively chemically inactive tin is greater than in zinc. Additionally, the sign of the response, whether positive or negative, can sometimes be influenced by the molecular state of the wire (see Chap. XII).
As regards the electrolyte, dilute NaCl solution, dilute solution of bichromate of potash &c. are normal in their action, that is to say, the electric response in such electrolytes is practically the same as with water. Ordinarily I use tap-water as the electrolyte. Zinc wires in ZnSO4 solution give responses similar in character to those given by, for example, Pt or Sn in water.
When it comes to the electrolyte, a diluted NaCl solution and a diluted solution of potassium bichromate, among others, work normally. This means that the electric response in these electrolytes is nearly the same as in water. Usually, I use tap water as the electrolyte. Zinc wires in a ZnSO4 solution generate responses similar to those produced by, for example, platinum or tin in water.
Test experiment.—It may be urged that the E.M. effect is due in some way (1) to the friction of the vibrating wire against the liquid; or (2) to some unknown surface action, at the point in the wire of the contact of liquid and air surfaces. This second objection has already been completely met in experi mental modification, fig. 55, b, where the wire was shown to give response when kept completely immersed in water, variation of surface being thus entirely eliminated.
Test experiment.—It's possible to argue that the E.M. effect is caused in some way (1) by the friction of the vibrating wire against the liquid; or (2) by some unknown surface interaction at the point where the wire meets the liquid and air surfaces. This second argument has already been fully addressed in experimental modification, fig. 55, b, where it was demonstrated that the wire responded while completely submerged in water, thereby eliminating any variations in surface.
Both these questions may, however, be subjected to a definite and final test. When the wire to be acted on is clamped below, and vibration is imparted to it, a strong molecular disturbance is produced. If now it be carefully released from the clamp, and the wire rotated backwards and forwards, there could be little molecular disturbance, but the liquid friction and surface variation, if any, would remain. The effect of any slight disturbance outstanding owing to shaking of the wire would be relatively very small.
Both of these questions can, however, be put to a clear and final test. When the wire to be acted on is clamped at the bottom and vibration is applied to it, a strong molecular disturbance occurs. If it is then carefully released from the clamp and the wire is rotated back and forth, there will be minimal molecular disturbance, but any liquid friction and surface variations, if present, will remain. The effect of any slight disturbance caused by shaking the wire will be relatively very small.
We can thus determine the effect of liquid friction and surface action by repeating an experiment with and without clamping. In a tin wire cell, with interposed external resistance equal to one million ohms, the wire A was subjected to a series of vibrations through 180°, and a deflection of 210 divisions was obtained. A corresponding negative deflection resulted on vibrating the wire B. Now A was released from the clamp, so that it could be rotated backwards and forwards in the water by means of the handle. On vibrating the wire A no measurable deflection was produced, thus showing that neither water friction nor surface variation had anything to do with the electric action. The vibration of the still clamped B gave rise to the normal strong deflection.
We can determine the effects of liquid friction and surface action by repeating an experiment with and without clamping. In a tin wire cell, with an external resistance of one million ohms, the wire A was subjected to a series of vibrations through 180°, resulting in a deflection of 210 divisions. A corresponding negative deflection occurred when vibrating the wire B. Then, A was released from the clamp, allowing it to be rotated back and forth in the water using a handle. When wire A was vibrated, no measurable deflection was produced, indicating that neither water friction nor surface variation played a role in the electric action. The vibration of the still clamped B resulted in the usual strong deflection.
A new and theoretically interesting molecular voltaic cell may thus be made, in which the two elements consist of the same metal. Molecular disturbance is in this case the main source of energy. A cell once made may be kept in working order for some time by pouring in a little vaseline to prevent evaporation of the liquid.
A new and theoretically fascinating molecular solar cell can be created, where both components are made of the same metal. In this scenario, molecular disruption is the primary source of energy. Once assembled, the cell can continue to function for a while if you pour in some vaseline to stop the liquid from evaporating.
It will be shown further, in succeeding chapters, by numerous instances, that any conditions which increase molecular mobility will also increase intensity of response, and conversely that any conditions having the reverse effect will depress response.
It will be demonstrated later, in upcoming chapters, through many examples, that any factors that enhance molecular movement will also boost the intensity of the response, and conversely, that any factors that have the opposite effect will reduce the response.
CHAPTER XII
INORGANIC RESPONSE—METHODS OF ENSURING CONSISTENT RESULTS
- Preparation of wire
- —Effect of single stimulus.
I shall now proceed to describe in detail the response-curves obtained with metals. The E.M. variations resulting from stimulus range, as has been said, from ·4 volt to ·01 of that value, according to the metal employed. And as these are molecular phenomena, the effect will also depend on the molecular condition of the wire.
I will now describe in detail the response curves obtained with metals. The E.M. variations resulting from the stimulus range, as mentioned, go from 0.4 volt to 0.01 of that value, depending on the metal used. And since these are molecular phenomena, the effect will also depend on the molecular condition of the wire.
Preparation of wire.—In order to have our results thoroughly consistent, it is necessary to bring the wire itself into a normal condition for experiment. The very fact of mounting it in the cell strains it, and the after-effect of this strain may cause irregularities in the response.
Preparation of wire.—To ensure our results are consistent, we need to adjust the wire to a normal condition for experimentation. Simply mounting it in the cell puts it under strain, and the lingering effects of that strain can lead to inconsistencies in the response.
For the purpose of bringing the wire to this normal state, one or all of the following devices may be used with advantage. (1) The wires obtained are usually wound on spools. It is, therefore, advisable to straighten any given length, before mounting, by holding it stretched, and rubbing it up and down with a piece of cloth. On washing with water, they are now ready for mounting in the cell.
To bring the wire back to its normal state, one or more of the following methods can be helpful. (1) The wires are typically wound on spools. It’s a good idea to straighten a specific length before mounting it by holding it taut and rubbing it up and down with a piece of cloth. After washing with water, they are now ready to be mounted in the cell.

Fig. 58.—Effect of Annealing on increasing the Response of both A and B Wires (Tin)
Fig. 58.—Impact of Annealing on Enhancing the Response of both A and B Wires (Tin)
Stimuli (vibration of 160°) applied at intervals of one minute.
Stimuli (vibration of 160°) applied every minute.
In this case it will be found helpful (3) to have recourse to the process of annealing. For if response be a molecular phenomenon, then anything that increases molecular mobility will also increase its intensity. Hence we may expect annealing to enhance responsiveness. This inference will be seen verified in the record given in fig. 58. In the case under consideration, the convenient method employed was by pouring hot water into the cell, and allowing it to stand and cool slowly. The first three pairs of responses were taken by stimulating A and B alternately, on mounting in the cell, which was filled with water. Hot water was then substituted, and the cell was allowed to cool down to its original temperature. The six following pairs of responses were then taken. That this beneficial effect of annealing was not due to any accidental circumstance will be seen from the fact that both wires have their sensitiveness equally enhanced.
In this case, it’s helpful to use the process of annealing. If response is a molecular phenomenon, then anything that increases molecular mobility will also boost its intensity. So, we can expect annealing to improve responsiveness. This conclusion is confirmed in the record given in fig. 58. For the situation we’re looking at, the method used was pouring hot water into the cell and letting it stand to cool slowly. The first three pairs of responses were taken by stimulating A and B alternately, while mounted in the cell filled with water. Hot water was then added, and the cell was allowed to cool down to its original temperature. The next six pairs of responses were then recorded. That the positive effect of annealing wasn’t due to some random circumstance is evident from the fact that both wires had their sensitivity equally enhanced.
(4) In addition to this mode of annealing, both wires may be short-circuited and vibrated for a time. Lastly (5) slight stretching in situ will also sometimes be found beneficial. For this purpose I have a screw arrangement.
(4) Along with this annealing method, both wires can be shorted and vibrated for a while. Finally, (5) slight stretching in situ can also be helpful at times. For this, I have a screw setup.
By one or all of these methods, with a little practice, it is always possible to bring the wires to a normal condition. The responses subsequently obtained become extraordinarily consistent. There is therefore no reason why perfect results should not be arrived at.
By one or all of these methods, with a little practice, it's always possible to get the wires back to a normal state. The responses you get afterward become really consistent. So, there’s no reason you shouldn't achieve perfect results.

Effect of single stimulus.—The accompanying figure (fig. 59) gives a series, each of which is the response curve for a single stimulus of uniform intensity, the amplitude of vibration being kept constant. The perfect regularity of responses will be noticed in this figure. The wire after a long period of rest may be in an abnormal condition, but after a short period of stimulation the responses become extremely regular, as may be noticed in this figure. Tin is, usually speaking, almost indefatigable, and I have often obtained several hundreds of successive responses showing practically no fatigue. In the figure it will be noticed that the rising portion of the curve is somewhat steep, and the recovery convex to the abscissa, the fall being relatively rapid in its first, and less rapid in its later, parts. As the electric variation is the concomitant effect of molecular disturbance—a temporary upset of the molecular equilibrium—on the cessation of the external stimulus, the excitatory state, and its expression in electric variation, disappear with the return of the molecules to their condition of equilibrium. This process is seen clearly in the curve of recovery.
Effect of single stimulus.—The figure (fig. 59) shows a series of response curves for a single stimulus at a consistent intensity, keeping the vibration amplitude constant. You can see the perfect regularity of these responses in this figure. After a long period of rest, the wire might be in an unusual state, but after a short period of stimulation, the responses quickly become very consistent, as shown in the figure. Tin is generally quite resilient, and I've often recorded several hundred successive responses with almost no signs of fatigue. In the figure, you’ll notice that the rising part of the curve is fairly steep, while the recovery part curves upward, with the decline happening relatively quickly at first and slowing down later on. When the external stimulus ends, the electric variation results from a temporary disturbance in the molecular equilibrium; as the molecules return to their equilibrium state, the excitatory condition and its electric expression fade away. This recovery process is clearly illustrated in the recovery curve.
Different metals exhibit different periods of recovery, and this again is modified by any influence which affects the molecular condition.
Different metals have varying recovery times, and this is also affected by any factors that influence their molecular state.
That the excitatory state persists for a time even on the cessation of stimulus can be independently shown by keeping the galvanometer circuit open during the application of stimulus, and completing it at various short intervals after the cessation, when a persisting electrical effect, diminishing rapidly with time, will be apparent. The rate of recovery immediately on the cessation of stimulus is rather rapid, but traces of strain persist for a short time.
The fact that the excitatory state continues for a while even after the stimulus stops can be demonstrated by keeping the galvanometer circuit open while applying the stimulus, and then closing it at various short intervals after the stimulus ends. This will show a lasting electrical effect that decreases quickly over time. The recovery rate right after the stimulus stops is fairly quick, but some traces of strain stick around for a short period.
CHAPTER XIII
INORGANIC RESPONSE—MOLECULAR MOBILITY: ITS INFLUENCE ON RESPONSE
- Effects of molecular inertia
- —Prolongation of period of recovery by overstrain
- —Molecular model
- —Reduction of molecular sluggishness attended by quickened recovery and heightened response
- —Effect of temperature
- —Modification of latent period and period of recovery by the action of chemical reagents
- —Diphasic variation.
We have seen that the stimulation of matter causes an electric variation, and that the acted substance gradually recovers from the effect of stimulus. We shall next study how the form of response-curves is modified by various agencies.
We have seen that stimulating matter causes an electric change, and that the affected substance slowly recovers from the impact of the stimulus. Next, we will examine how different factors alter the shape of response curves.
In order to study these effects we must use, in practice, a highly sensitive galvanometer as the recorder of E.M. variations. This necessitates the use of an instrument with a comparatively long period of swing of needle, or of suspended coil (as in a D’Arsonval). Owing to inertia of the recording galvanometer, however, there is a lag produced in the records of E.M. changes. But this can be distinguished from the effect of the molecular inertia of the substance itself by comparing two successive records taken with the same instrument, in one of which the latter effect is relatively absent, and in the other present. We wish, for example, to find out whether the E.M. effect of mechanical stimulus is instantaneous, or, again, whether the effect disappears immediately. We first take a galvanometer record of the sudden introduction and cessation of an E.M.F. on the circuit containing the vibration-cell (fig. 60, a). We then take a record of the E.M. effect produced by a stimulus caused by a single torsional vibration. In order to make the conditions of the two experiments as similar as possible, the disturbing E.M.F., from a potentiometer, is previously adjusted to give a deflection nearly equal to that caused by stimulus. The torsional vibration was accomplished in a quarter of a second, and the contact with the potentiometer circuit was also made for the same length of time.
To study these effects, we need to use a highly sensitive galvanometer to record E.M. variations. This requires an instrument with a relatively long needle swing or suspended coil (like in a D’Arsonval). However, due to the inertia of the recording galvanometer, there's a delay in the records of E.M. changes. We can differentiate this from the molecular inertia of the substance itself by comparing two consecutive records taken with the same instrument, where one shows the latter effect relatively absent, and the other shows it present. For example, we want to find out whether the E.M. effect of mechanical stimulus is instantaneous or if the effect disappears right away. First, we take a galvanometer record of the sudden introduction and cessation of an E.M.F. in the circuit containing the vibration-cell (fig. 60, a). Then we record the E.M. effect caused by a single torsional vibration. To make the conditions of the two experiments as similar as possible, we previously adjust the disturbing E.M.F. from a potentiometer to give a deflection almost equal to that caused by the stimulus. The torsional vibration was completed in a quarter of a second, and the contact with the potentiometer circuit was also made for the same duration.

Fig. 60
Fig. 60
(a) Arrangement for applying a short-lived E.M.F.
(b) Difference in the periods of recovery: (1) from instantaneous
E.M.F.; and (2) that caused by mechanical stimulus.
(a) Setup for applying a temporary E.M.F.
(b) Difference in recovery times: (1) from instant E.M.F.; and (2) caused by mechanical stimulus.
The record was then taken as follows. The recording drum had a fast speed of six inches in a minute, one of the small subdivisions representing a second. The battery contact in the main potentiometer circuit was made for a quarter of a second as just mentioned and a record taken of the effect of a short-lived E.M.F. on the circuit containing the cell. (2) A record was next taken of the E.M. variation produced in the cell by a single stimulus. It will be seen on comparison of the two records that the maximum effect took place relatively later in the case of mechanical stimulus, and that whereas the galvanometer recovery in the former case took place in 11 seconds, the recovery in the latter was not complete till after 60 seconds (fig. 60, b). This shows that it takes some time for the effect of stimulus to attain its maximum, and that the effect does not disappear till after the lapse of a certain interval. The time of recovery from strain depends on the intensity of stimulus. It takes a longer time to recover from a stronger stimulus. But, other things being equal, successive recovery periods from successive stimulations of equal intensity are, generally speaking, the same.
The record was taken as follows. The recording drum had a fast speed of six inches per minute, with one of the small subdivisions representing a second. The battery contact in the main potentiometer circuit was made for a quarter of a second, as mentioned earlier, and a record was made of the effect of a short-lived E.M.F. on the circuit containing the cell. (2) Next, a record was taken of the E.M. variation produced in the cell by a single stimulus. Comparing the two records, you'll see that the maximum effect occurred relatively later with a mechanical stimulus. The galvanometer recovery in the former case took 11 seconds, while the recovery in the latter wasn’t complete until after 60 seconds (fig. 60, b). This shows that it takes time for the effect of a stimulus to reach its maximum, and that the effect doesn't disappear until after a certain interval. The recovery time from strain depends on the intensity of the stimulus. It takes longer to recover from a stronger stimulus. However, all else being equal, the recovery periods from successive stimulations of the same intensity are generally consistent.
We may now study the influence of any change in external conditions by observing the modifications it produces in the normal curve.
We can now examine how any change in external conditions affects the normal curve by observing the changes it creates.

Fig. 61.—Prolongation of Period of Recovery after Overstrain
Fig. 61.—Extension of Recovery Period after Overexertion
Recovery is complete in 60″ when the stimulus is due to 20° vibration. But with stronger stimulus of 40° vibration, the period of recovery is prolonged to 90″.
Recovery is complete in 60 seconds when the stimulus is due to 20° vibration. But with a stronger stimulus of 40° vibration, the recovery period is extended to 90 seconds.
Prolongation of period of recovery by overstrain.—The pair of records given in fig. 61 shows how recovery is delayed, as the effect of overstrain. Curve (a) is for a single stimulus due to a vibration of amplitude 20°, and curve (b) for a stimulus of 40° amplitude of vibration. It will be noticed how relatively prolonged is the recovery in the latter case.
Extended Recovery Time Due to Overexertion.—The pair of records shown in fig. 61 illustrates how recovery is delayed because of overexertion. Curve (a) represents a single stimulus caused by a vibration with an amplitude of 20°, while curve (b) corresponds to a stimulus with a vibration amplitude of 40°. It's noticeable how much longer the recovery takes in the second case.

Molecular Model.—We have seen that the electric response is an outward expression of the molecular disturbance produced by the action of the stimulus. The rising part of the response-curve thus exhibits the effect of molecular upset, and the falling part, or recovery, the restoration to equilibrium. The mechanical model (fig. 62) will help us to visualise many complex response phenomena. The molecular model consists of a torsional pendulum—a wire with a dependent sphere. By the stimulus of a blow there is produced a torsional vibration—a response followed by recovery. The writing lever attached to the pendulum records the response-curves. The form of these curves, stimulus remaining constant, will be modified by friction; the less the friction, the greater is the mobility. The friction may be varied by more or less raising a vessel of sand touching the pendulum. By varying the friction the following curves were obtained.
Molecular Model.—We've established that the electric response is a visible sign of the molecular disturbance caused by the stimulus. The rising section of the response curve shows the impact of this molecular disruption, while the falling section, or recovery, indicates the return to balance. The mechanical model (fig. 62) will help us understand many complex response phenomena. The molecular model features a torsional pendulum—a wire with a hanging sphere. When struck, it creates a torsional vibration—a response followed by recovery. The writing lever connected to the pendulum records the response curves. The shape of these curves, with the stimulus held constant, will be affected by friction; less friction leads to greater mobility. The friction can be adjusted by lifting a vessel of sand that makes contact with the pendulum. By changing the friction, the following curves were obtained.
(a) When there is little friction we get an after-oscillation, to which we have the corresponding phenomenon in the retinal after-oscillation (compare fig. 105).
(a) When there is little friction, we experience an after-oscillation, which corresponds to the retinal after-oscillation (see fig. 105).
(b and c) If the friction is increased, there is a damping of oscillation. In (c) we get recovery-curves similar to those found in nerve, muscle, plant, and metal.
(b and c) If friction increases, oscillation is dampened. In (c), we observe recovery curves similar to those seen in nerves, muscles, plants, and metals.
(d) If the friction is still further increased the maximum is reached much later, as will be seen in the increasing slant of the rising part of the curve; the height of response is diminished and the period of recovery very much prolonged by partial molecular arrest. The curve (d) is very similar to the ‘molecular arrest’ curve obtained by small dose of chemical reagents which act as ‘poison’ on living tissue or on metals (compare fig. 93, a).
(d) If the friction is increased even more, the peak happens much later, as shown by the increasing slope of the upward part of the curve; the level of response is lower, and the recovery time is greatly extended due to partial molecular arrest. The curve (d) looks a lot like the ‘molecular arrest’ curve obtained from a small dose of chemical agents that act as ‘poisons’ on living tissue or metals (compare fig. 93, a).
(e) When the molecular mobility is further decreased there is no recovery (compare fig. 93, b).
(e) When the molecular mobility decreases even more, there is no recovery (compare fig. 93, b).
Still further increase of friction completely arrests the molecular pendulum, and there is no response.
Still more friction completely stops the molecular pendulum, and there is no movement.
Reduction of molecular sluggishness attended (1) by quickened recovery.—Sometimes, after a cell has been resting for too long a period, especially on cold days, the wire gets into a sluggish condition, and the period of recovery is thereby prolonged. But successive vibrations gradually remove this inertness, and recovery is then hastened. This is shown in the accompanying curves, fig. 63, where (a) exhibits only very partial recovery even after the expiration of 60 seconds, whereas when a few vibrations had been given recovery was entirely completed in 47 seconds (b). There was here little change in the height of response.
Reduction of molecular sluggishness attended (1) by quicker recovery.—Sometimes, after a cell has been resting for too long, especially on cold days, the wire becomes sluggish, and the recovery period is extended. However, repeated vibrations gradually eliminate this sluggishness, allowing for faster recovery. This is illustrated in the accompanying curves, fig. 63, where (a) shows only minimal recovery even after 60 seconds, while with a few vibrations given, recovery was fully achieved in 47 seconds (b). There was little change in the height of response.

Fig. 63
Fig. 63
(a) Slow recovery of a wire in a sluggish condition.
(b) Quickened recovery in the same wire after a few vibrations.
(a) Slow recovery of a wire in a sluggish state.
(b) Faster recovery in the same wire after a few vibrations.
Or (2) by heightened response.—The removal of sluggishness by vibration, resulting in increased molecular mobility, is in other instances attended by increase in the height of response, as will be seen from the two sets of records which follow (fig. 64). Cold, due to prevailing frosty weather, had made the wires in the cell somewhat lethargic. The records in (a) were the first taken on the day of the experiment. The amplitudes of vibration were 45°, 90°, and 135°. In (b) are given the records of the next series, which are in every case greater than those of (a). This shows that previous vibration, by conferring increased mobility, had heightened the response. In this case, removal of molecular sluggishness is attended by greater intensity of response, without much change in the period of recovery. In connection with this it must be remembered that greater strain consequent on heightened response has a general tendency to a prolongation of the period of recovery.
Or (2) by increased response.—The removal of sluggishness through vibration, which leads to greater molecular mobility, is sometimes accompanied by an increase in response height, as shown in the two sets of records that follow (fig. 64). The cold weather had made the wires in the cell somewhat sluggish. The records in (a) were the first taken on the day of the experiment. The vibration amplitudes were 45°, 90°, and 135°. In (b), the records from the next series are all higher than those in (a). This indicates that the earlier vibration, by increasing mobility, had enhanced the response. In this case, the reduction of molecular sluggishness is accompanied by a greater intensity of response, with little change in the recovery period. It should be noted that greater strain resulting from heightened response generally tends to lengthen the recovery period.

Fig. 64
Fig. 64
(a) Three sets of responses for 45°, 90°, and 135° vibration in a
sluggish wire.
(b)The next three sets of responses in the same wire; increased
mobility conferred by previous vibration has heightened the response.
(a) Three sets of responses for 45°, 90°, and 135° vibration in a slow wire.
(b) The next three sets of responses in the same wire; the increased mobility from earlier vibrations has improved the response.
It is thus seen that when the wire is in a sluggish condition, successive vibrations confer increased molecular mobility, which finds expression in quickened recovery or heightened response.
It’s clear that when the wire is in a slow state, repeated vibrations enhance molecular movement, resulting in faster recovery or a stronger response.
Effect of temperature.—Similar considerations lead us to expect that a moderate rise of temperature will be conducive to increase of response. This is exhibited in the next series of records. The wire at the low temperature of 5° C. happened to be in a sluggish condition, and the responses to vibrations of 45° to 90° in amplitude were feeble. Tepid water at 30° C. was now substituted for the cold water in the cell, and the responses underwent a remarkable enhancement. But the excessive molecular disturbance caused by the high temperature of 90° C. produced a great diminution of response (fig. 65).
Effect of temperature.—Similar thoughts lead us to expect that a moderate increase in temperature will result in a greater response. This is shown in the next set of records. The wire at the low temperature of 5° C. was in a sluggish state, and the responses to vibrations with amplitudes from 45° to 90° were weak. Tepid water at 30° C. was then used instead of the cold water in the cell, and the responses showed a remarkable improvement. However, the excessive molecular disturbance caused by the high temperature of 90° C. resulted in a significant decrease in response (fig. 65).

Fig. 65.—Responses of a Wire To Amplitudes of Vibration 45° and 90°
Fig. 65.—Responses of a Wire to Vibration Amplitudes at 45° and 90°
(a) Responses when the wire was in a sluggish condition at temperature
of 5° C.
(b) Enhanced response at 30° C.
(c) Diminution of response at 90° C.
(a) Responses when the wire was in a slow state at a temperature of 5° C.
(b) Improved response at 30° C.
(c) Decrease in response at 90° C.
Diphasic variation.—It has already been said that if two points A and B are in the same physico-chemical condition, then a given stimulus will give rise to similar excitatory electric effects at the two points. If the galvanometer deflection is ‘up’ when A alone is excited, the excitation of B will give rise to a downward deflection. When the two points are simultaneously excited the electric variation at the two points will continuously balance each other. Under such conditions there will be no resultant deflection. But if the intensity of stimulation of one point is relatively stronger, then the balance will be disturbed, and a resultant deflection produced whose sign and magnitude can be found independently by the algebraical summation of the individual effects of A and B.
Diphasic variation.—It has already been established that if two points A and B are in the same physical and chemical condition, then a specific stimulus will generate similar excitatory electric effects at both points. If the galvanometer shows a deflection ‘up’ when only A is stimulated, then stimulating B will result in a downward deflection. When both points are excited at the same time, the electric variations at both points will continuously counterbalance each other. In this case, there will be no overall deflection. However, if the stimulation intensity at one point is relatively stronger, the balance will be disrupted, resulting in a deflection whose direction and size can be determined by simply adding together the individual effects of A and B.
It has also been shown that a balancing point for the block, which is approximately near the middle of the wire, may be found so that the vibrations of A and B through the same amplitude produce equal and opposite deflection. Simultaneous vibration of both will give no resultant current; when the block is abolished and the wire is vibrated as a whole, there will still be no resultant, inasmuch as similar excitations are produced at A and B.
It has also been shown that a balance point for the block, which is roughly located near the middle of the wire, can be found so that the vibrations of A and B with the same amplitude produce equal and opposite deflections. If both vibrate at the same time, there will be no resultant current; when the block is removed and the wire is vibrated as a whole, there will still be no resultant since similar excitations occur at A and B.
After obtaining the balance, if we apply an exciting reagent like Na2CO3 at one point, and a depressing reagent like KBr at the other, the responses will now become unequal, the more excitable point giving a stronger deflection. We can, however, make the two deflections equal by increasing the amplitude of vibration of the less sensitive point. The two deflections may thus be rendered equal and opposite, but the time relations—the latent period, the time rate for attaining the maximum excitation and recovery from that effect—will no longer be the same in the two cases. There would therefore be no continuous balance, and we obtain instead a very interesting diphasic record. I give below an exact reproduction of the response-curves of A and B recorded on a fast-moving drum. It will be remembered that one point was touched with Na2CO3 and the other with KBr. By suitably increasing the amplitude of vibration of the less sensitive, the two deflections were rendered approximately equal. The records of A and B were at first taken separately (fig. 66, a). It will be noticed that the maximum deflection of A was attained relatively much earlier than that of B. The resultant curve R′ was obtained by summation.
After getting the balance, if we apply an exciting reagent like Na2CO3 on one side and a depressing reagent like KBr on the other, the responses will become unequal, with the more reactive side showing a stronger deflection. However, we can make the two deflections equal by increasing the vibration amplitude of the less sensitive side. The two deflections can be made roughly equal and opposite, but the timing—like the latent period, the time taken to reach maximum excitation, and the recovery time—won't be the same in both cases. Because of this, there won’t be a continuous balance, and we end up with a very interesting diphasic record. Below is an exact reproduction of the response curves of A and B recorded on a fast-moving drum. Remember that one side was treated with Na2CO3 and the other with KBr. By appropriately increasing the vibration amplitude of the less sensitive side, the two deflections were made approximately equal. The records of A and B were initially taken separately (fig. 66, a). It's noticeable that the maximum deflection of A was reached significantly earlier than that of B. The resulting curve R' was created by summation.

Fig. 66.—Diphasic Variation
Fig. 66.—Diphasic Variation
(a) Records of A and B obtained separately. R′ is the resultant by algebraical summation. (b) Diphasic record obtained by simultaneous stimulation of A and B.
(a) Records of A and B obtained separately. R′ is the result from adding them together algebraically. (b) Diphasic record obtained from stimulating A and B at the same time.
After taking the records of A and B separately, a record of resultant effect R due to simultaneous vibration of A and B was next taken. It gave the curious two-phased response—positive effect followed by negative after-vibration, practically similar to the resultant curve R′ (fig. 66, b).
After recording the effects of A and B individually, a record of the combined effect R due to the simultaneous vibrations of A and B was then taken. It produced an interesting two-phased response—first a positive effect followed by a negative after-vibration, which was practically similar to the resultant curve R′ (fig. 66, b).
The positive portion of the curve is due to A effect and the negative to B. If by any means, say by either increasing the amplitude of vibration of A or increasing its sensitiveness, the response of A is very greatly enhanced, then the positive effect would be predominant and the negative effect would become inconspicuous. When the two constituent responses are of the same order of magnitude, we shall have a positive response followed by a negative after-vibration; the first twitch will belong to the one which responds earlier. If the response of A is very much reduced, then the positive effect will be reduced to a mere twitch and the negative effect will become predominant.
The positive part of the curve is due to A effect and the negative part is due to B. If, for any reason—like increasing the vibration amplitude of A or boosting its sensitivity—the response from A is significantly increased, then the positive effect would take over and the negative effect would be less noticeable. When both responses are similar in strength, we’ll see a positive response followed by a negative after-effect; the first movement will come from the one that reacts sooner. If the response from A is significantly weakened, then the positive effect will shrink to just a slight movement, and the negative effect will dominate.
I give a series of records, fig. 67, in which these three principal types are well exhibited, the two contacts having been rendered unequally excitable by solutions of the two reagents KBr and Na2CO3. A and B were vibrated simultaneously and records taken. (a) First, the relative response of B (downward) is increased by increasing its amplitude of vibration. The amplitude of vibration of A was throughout maintained constant. The negative or downward response is now very conspicuous, there being only a mere preliminary indication of the positive effect. (b) The amplitude of vibration of B is now slightly reduced, and we obtain the diphasic effect. (c) The intensity of vibration of B is diminished still further, and the negative effect is seen reduced to a slight downward after-vibration, the positive up-curve being now very prominent (fig. 67).
I provide a series of records, fig. 67, that clearly show these three main types. The two contacts have been made unevenly responsive by solutions of the two reagents KBr and Na2CO3. A and B were vibrated at the same time, and records were taken. (a) First, the relative response of B (downward) increases as its vibration amplitude is raised. The vibration amplitude of A was kept constant throughout. The negative or downward response is very noticeable now, with only a slight initial indication of the positive effect. (b) Now, the vibration amplitude of B is slightly reduced, and we see the diphasic effect. (c) The vibration intensity of B is further decreased, and the negative effect becomes a slight downward after-vibration, while the positive upward curve is now very prominent (fig. 67).

Fig. 67.—Negative, Diphasic, and Positive Resultant Response
Fig. 67.—Negative, Diphasic, and Positive Resultant Response
Continuous transformation from negative to positive.—I have shown the three phases of transformation, the intensity of one of the constituent responses being varied by altering the intensity of disturbance.
Continuous change from negative to positive.—I have demonstrated the three stages of transformation, with the strength of one of the component responses being adjusted by changing the strength of the disturbance.
In the following record (fig. 68) I succeeded in obtaining a continuous transformation from positive to negative phase by a continuous change in the relative sensitiveness of the two contacts.
In the following record (fig. 68), I managed to achieve a smooth transition from positive to negative phase by continuously adjusting the relative sensitivity of the two contacts.
I found that traces of after-effect due to the application of Na2CO3 remain for a time. If the reagent is previously applied to an area and the traces of the carbonate then washed off, the increased sensitiveness conferred disappears gradually. Again, if we apply Na2CO3 solution to a fresh point, the sensitiveness gradually increases. There is another further interesting point to be noticed: the beginning of response is earlier when the application of Na2CO3 is fresh.
I found that there are lingering effects from using Na2CO3 that last for a while. If the reagent is applied to an area and then the leftover carbonate is washed away, the increased sensitivity wears off gradually. On the other hand, if we apply the Na2CO3 solution to a new spot, the sensitivity gradually increases again. There's one more interesting point to note: the response starts sooner when the Na2CO3 is freshly applied.

Fig. 68.—Continuous Transformation from Negative To Positive through Intermediate Diphasic Response
Fig. 68.—Ongoing Change from Negative to Positive through Intermediate Two-Phase Response
Thick dots represent the times of application of successive stimuli.
Thick dots indicate when each stimulus was applied.
We have thus a wire held at one end, and successive uniform vibrations at intervals of one minute imparted to the wire as a whole, by means of a vibration head on the other end.
We have a wire that's secured at one end, with consistent uniform vibrations applied to the entire wire at one-minute intervals, using a vibration head on the opposite end.
Owing to the after-effect of previous application of Na2CO3 the sensitiveness of B is at the beginning great, hence the three resultant responses at the beginning are negative or downward.
Due to the after-effect of the earlier application of Na2CO3, the sensitivity of B is initially high, which is why the first three responses are negative or downward.
Dilute solution of Na2CO3 is next applied to A. The response of A (up) begins earlier and continues to grow stronger and stronger. Hence, after this application, the response shows a preliminary positive twitch of A followed by negative deflection of B. The positive grows continuously. At the fifth response the two phases, positive and negative, become equal, after that the positive becomes very prominent, the negative being reduced as a feeble after-vibration.
A dilute solution of Na2CO3 is then applied to A. The response of A (up) starts sooner and continues to strengthen. As a result, after this application, the response shows an initial positive twitch of A, followed by a negative deflection of B. The positive response keeps growing continuously. At the fifth response, the two phases, positive and negative, become equal; after that, the positive becomes very noticeable, while the negative fades into a weak after-vibration.
It need only be added here that the diphasic variations as exhibited by metals are in every way counterparts of similar phenomena observed in animal tissues.
It should just be noted here that the diphasic variations shown by metals are in every way similar to the phenomena observed in animal tissues.
CHAPTER XIV
INORGANIC RESPONSE—FATIGUE, STAIRCASE, AND MODIFIED RESPONSE
- Fatigue in metals
- —Fatigue under continuous stimulation
- —Staircase effect
- —Reversed responses due to molecular modification in nerve and metal, and their transformation into normal after continuous stimulation
- —Increased response after continuous stimulation.

Fatigue.—In some metals, as in muscle and in plant, we find instances of that progressive diminution of response which is known as fatigue (fig. 69). The accompanying record shows this in platinum (fig. 70). It has been said that tin is practically indefatigable. We must, however, remember that this is a question of degree only. Nothing is absolutely indefatigable. The exhibition of fatigue depends on various conditions. Even in tin, then, I obtained the characteristic fatigue-curve with a specimen which had been in continuous use for many days (fig. 71). While discussing the subject of fatigue in plants, I have adduced considerations which showed that the residual effect of strain was one of the main causes for the production of fatigue. This conclusion receives independent support from the records obtained with metals.
Fatigue.—In some metals, like in muscle and plants, we see examples of the gradual reduction in response known as fatigue (fig. 69). The accompanying record demonstrates this in platinum (fig. 70). It's been said that tin is almost tireless. However, we should remember that this is only a matter of degree. Nothing is completely tireless. The manifestation of fatigue depends on various factors. Even in tin, I obtained the typical fatigue curve from a specimen that had been in continuous use for several days (fig. 71). While discussing fatigue in plants, I presented considerations that showed the lingering effects of strain were one of the main causes of fatigue. This conclusion is further supported by the data obtained with metals.


Fig. 71.—Fatigue Shown by Tin Wire which had been Continuously Stimulated for Several Days
Fig. 71.—Fatigue Shown by Tin Wire that had been Continuously Stimulated for Several Days
In this connection the important fact is that the various typical fatigue effects exhibited in living substances are exactly reproduced in metals, where there can be question neither of fatigue-product producing fatigue effects, nor of those constructive processes by which they might be removed. We have seen, both in muscles and in plants, that if sufficient time for complete recovery be allowed between each pair of stimuli, the heights of successive responses are the same, and there is no apparent fatigue (see page 39). But the height of response diminishes as the excitation interval is shortened. We find the same thing in metals. Below is given a record taken with tin (fig. 72). Throughout the experiment the amplitude of vibration was maintained constant, but in (a) the interval between consecutive stimuli was 1′, while in (b) this was reduced to 30″. A diminution of height immediately occurs. On restoring the original rhythm as in (c), the responses revert to their first large value. Thus we see that when the wire has not completely recovered, its responses, owing to residual strain, undergo diminution. Height of response is thus decreased by incomplete recovery. If then sufficient time be not allowed for perfect recovery, we can understand how, under certain circumstances, the residual strain would progressively increase with repetition of stimulus, and thus there would be a progressive diminution of height of response or fatigue. Again, we saw in the last chapter that increase of strain necessitates a longer period of recovery. Thus the longer a wire is stimulated, the more and more overstrained it becomes, and it therefore requires a gradual prolongation of the interval between the successive stimuli, if recovery is to be complete. This interval, however, being maintained constant, the recovery periods virtually undergo a gradual reduction, and successive recoveries become more and more incomplete. These considerations may be found to afford an insight into the progressive diminution of response in fatigued substances.
In this context, the key point is that the various typical fatigue effects seen in living substances are exactly replicated in metals, where there is no question of fatigue products causing fatigue effects, nor of any processes that could remove them. We have observed in both muscles and plants that if enough time is allowed for complete recovery between each pair of stimuli, the heights of successive responses remain the same, and there is no visible fatigue (see page 39). However, as the time interval between stimuli is shortened, the height of the response decreases. The same phenomenon occurs in metals. Below is a record taken with tin (fig. 72). Throughout the experiment, the amplitude of vibration stayed constant, but in (a) the interval between consecutive stimuli was 1 minute, whereas in (b) it was reduced to 30 seconds. A decrease in height occurs immediately. When the original rhythm is restored as in (c), the responses return to their original high value. Thus, when the wire has not fully recovered, its responses diminish due to residual strain. The height of the response decreases because of incomplete recovery. If sufficient time is not allowed for complete recovery, we can see how, under certain conditions, the residual strain would progressively increase with repetitive stimuli, resulting in a progressive decrease in the response height, or fatigue. Additionally, we noted in the last chapter that an increase in strain requires a longer recovery period. Thus, the longer a wire is stimulated, the more overstrained it becomes, necessitating a gradual extension of the interval between successive stimuli for full recovery. However, if this interval remains constant, the recovery periods effectively undergo a gradual reduction, leading to increasingly incomplete successive recoveries. These considerations provide insight into the progressive decrease of response in fatigued substances.

Fig. 72.—Diminution of Response due to Shortening the Period of Recovery
Fig. 72.—Reduced Response from Shortening the Recovery Time
The stimulus is maintained constant. In (a) the interval between two successive stimuli is one minute, in (b) it is half a minute, and in (c) it is again one minute. The response in (b) is feebler than in either (a) or (c).
The stimulus stays the same. In (a), the gap between two successive stimuli is one minute; in (b), it's half a minute; and in (c), it's back to one minute. The response in (b) is weaker than in both (a) and (c).
Fatigue under continuous stimulation.—Fatigue is perhaps best shown under continuous stimulation. For example, in muscles, when fresh and not fatigued, the top of the tetanic curve is horizontal, or may even be ascending, but with long-continued stimulation the curve declines. The rapidity of this decline depends on the nature of the muscle and its previous condition.
Fatigue under continuous stimulation.—Fatigue is probably most clearly demonstrated under constant stimulation. For instance, in muscles, when they are fresh and not fatigued, the peak of the tetanic curve is flat or may even be rising, but with prolonged stimulation, the curve drops. The speed of this decline depends on the type of muscle and its previous state.
In metals I have found exactly parallel instances. In tin, so little liable to fatigue, the top of the curve is horizontal or ascending; or it may exhibit a slight decline. But the record with platinum shows the rapid decline due to fatigue (fig. 73).
In metals, I've found similar examples. In tin, which is not very prone to fatigue, the top of the curve is flat or rising; it might also show a slight drop. But the data for platinum shows a sharp decline due to fatigue (fig. 73).

Fig. 73
Fig. 73
(a) The top of response-curve under continuous stimulation in tin
is horizontal or ascending as in figure.
(b) In platinum there is
rapid decline owing to fatigue.
(a) The peak of the response curve under constant stimulation in tin is flat or rising, as shown in the figure.
(b) In platinum, there is a quick drop due to fatigue.
Taking any of these instances, say that in which fatigue is most prominent, it is found that short period of rest restores the original intensity of response. This affords additional proof of the fact that fatigue is due to overstrain, and that this strain, with its sign of attendant fatigue, disappears with time.
Taking any of these examples, like the one where fatigue is most evident, we see that a short break restores the original intensity of response. This provides further evidence that fatigue is caused by overexertion, and that this strain, along with its associated fatigue, fades over time.
Staircase effect.—We shall now discuss an effect which appears to be the direct opposite of fatigue. This is the curious phenomenon known to physiologists as ‘the staircase’ effect, in which successive uniform stimuli produce a series of increasing responses. This is seen under particular conditions in the response of certain muscles (fig. 74, a). It is also observed sometimes even in nerve, which otherwise, generally speaking, gives uniform responses. Of this effect, no satisfactory theory has as yet been offered. It is in direct contradiction to that theory which supposes that each stimulus is followed by dissimilation or break-down of the tissue, reducing its function below par. For in these cases the supposed dissimilation is followed not by a decrease but by an increase of functional activity. This ‘staircase effect’ I have shown to be occasionally exhibited by plants. I have also found it in metals. In the last chapter we have seen that a wire often falls, especially after resting for a long time, into a state of comparative sluggishness, and that this molecular inertness then gradually gives place to increased mobility under stimulation. As a consequence, an increased response is thus obtained. I give in fig. 74, b, a series of responses to uniform stimuli, exhibited by platinum which had been at rest for some time. This effect is very clearly shown here. So we see that in a substance which has previously been in a sluggish condition, stimulation confers increased mobility. Response thus reaches a maximum, but continued stimulation may afterwards produce overstrain, and the subsequent responses may then show a decline. This consideration will explain certain types of responses exhibited by muscles, where the first part of the series exhibits a staircase increase followed by declining responses of fatigue.
Staircase effect.—Now, let's talk about an effect that seems to be the exact opposite of fatigue. This is the interesting phenomenon known to physiologists as the "staircase" effect, where a series of consistent stimuli lead to progressively stronger responses. This can be seen under specific conditions in the reactions of certain muscles (fig. 74, a). It's also sometimes noted in nerves, which generally produce uniform responses. No satisfactory theory has been proposed for this effect yet. It directly contradicts the theory that each stimulus leads to the breakdown or disintegration of tissue, which would lower its functionality. In these cases, the expected breakdown is followed not by a decrease but by an increase in functional activity. I have shown that this "staircase effect" is occasionally displayed by plants, and I have also found it in metals. In the last chapter, we noted that a wire often becomes sluggish, especially after being at rest for a long time, and this molecular inactivity gradually shifts to increased mobility when stimulated. Consequently, an increased response is achieved. I present in fig. 74, b, a series of responses to consistent stimuli from platinum that had been at rest for a while. This effect is very clearly demonstrated here. Thus, we see that in a substance that has been in a sluggish state, stimulation results in greater mobility. Responses reach a peak, but continued stimulation can cause overstrain, leading to a decline in subsequent responses. This explains certain response patterns seen in muscles, where the initial part of the sequence shows a staircase increase followed by decreasing responses due to fatigue.

Fig. 74.—‘Staircase’ Effect
Fig. 74.—‘Staircase’ Effect
(a) in muscle (after Engelmann).
(b) in metal.
(a) in muscle (after Engelmann).
(b) in metal.
Reversed response due to molecular modification and its transformation into normal after continuous stimulation (1) in nerve.—Reference has already been made to the fact that a nerve which, when fresh, exhibited the normal negative response, will often, if kept for some time in preservative saline, undergo a molecular modification, after which it gives a positive variation. Thus while the response given by fresh nerve is normal or negative, a stale nerve gives modified, i.e. reversed or positive, response. This peculiar modification does not always occur, yet is too frequent to be considered abnormal. Again, when such a nerve is subjected to tetanisation or continuous stimulation, this modified response tends once more to become normal.
Reversed response due to molecular changes and its return to normal after constant stimulation (1) in nerve.—It has already been noted that a nerve that initially shows a normal negative response when fresh will often, if stored for a while in preservative saline, undergo molecular changes, resulting in a positive variation. Therefore, while the response of fresh nerve is normal or negative, a stale nerve exhibits a modified, meaning reversed or positive, response. This unusual modification doesn’t always happen, but it occurs frequently enough to not be considered abnormal. Furthermore, when such a nerve is subjected to tetanization or continuous stimulation, this modified response tends to revert back to normal.
It is found that not only tetanisation, but also CO2 has the power of converting the modified response into normal. Hence it has been suggested that the conversion under tetanisation of modified response to normal, in stale nerve, is due to a hypothetical evolution of CO2 in the nerve during stimulation.[16]
It’s found that not only tetanization, but also CO2, can change the modified response back to normal. Therefore, it’s suggested that the change from a modified response to normal during tetanization in a stale nerve is due to a hypothetical buildup of CO2 in the nerve during stimulation.[16]
(2) In metals.—I have, however, met with exactly parallel phenomena in metals, where, owing to some molecular modification, the responses became reversed, and where, under continuous stimulation, though here there could be no possibility of the evolution of CO2, they tended again to become normal.
(2) In metals.—I have, however, encountered exactly similar phenomena in metals, where, due to some molecular changes, the responses were inverted, and where, under constant stimulation, even though there was no chance of CO2 being produced, they tended to return to a normal state.
If after mounting a wire in a cell filled with water, it be set aside for too long a time, I have sometimes noticed that it undergoes a certain modification, owing to which its response ceases to be normal and becomes reversed in sign. I have obtained this effect with various metals, for instance lead and tin, and even with the chemically inactive substance—platinum.
If you mount a wire in a cell filled with water and leave it sitting for too long, I've noticed that it changes in a way that causes its response to become abnormal and reversed. I've seen this happen with different metals, like lead and tin, and even with the chemically stable substance—platinum.

Fig. 75.—Abnormal Positive (up) Response in Nerve Converted into Normal (down) Response after Continuous Stimulation T (Waller)
Fig. 75.—Abnormal Positive (up) Response in Nerve Converted into Normal (down) Response after Continuous Stimulation T (Waller)
The galvanometer is not dead-beat, and shows after-oscillation.
The galvanometer isn’t dead-beat and shows after-oscillation.
The subject will be made clearer if we first follow in detail the phenomenon exhibited by modified nerve, giving this abnormal response. The normal responses in nerve are usually represented by ‘down’ and the reversed abnormal responses by ‘up’ curves. In the modified nerve, then, the abnormal responses are ‘up’ instead of the normal ‘down.’ The record of such abnormal response in the modified nerve is shown in fig. 75. It will be noticed that in this, the successive responses are undergoing a diminution, or tending towards the normal. After continuous stimulation or tetanisation (T), it will be seen that the abnormal or ‘up’ responses are converted into normal or ‘down.’
The topic will become clearer if we first look closely at the phenomenon shown by modified nerves, which give this abnormal response. Normal nerve responses are usually shown as 'down' curves, while the reversed abnormal responses are shown as 'up' curves. In the modified nerve, the abnormal responses appear as 'up' instead of the normal 'down.' The record of this abnormal response in the modified nerve is shown in fig. 75. You will notice that in this case, the successive responses are decreasing or moving towards normal. After continuous stimulation or tetanization (T), it becomes clear that the abnormal or 'up' responses change into normal or 'down.'
I shall now give a record which will exhibit an exactly similar transformation from the abnormal to normal response after continuous stimulation. Here the normal responses are represented by ‘up’ and the abnormal by ‘down’ curves. This record was given by a tin wire, which had been molecularly modified (fig. 76). We have at first the abnormal responses; successive responses are undergoing a diminution or tending towards the normal; after continuous stimulation (T), the subsequent responses are seen to have become normal. Another record, obtained with platinum, shows the same phenomenon (fig. 77).
I’m now going to provide a record that shows a similar change from abnormal to normal responses after continuous stimulation. In this case, the normal responses are represented by ‘up’ curves and the abnormal by ‘down’ curves. This record was produced by a tin wire that had been molecularly modified (fig. 76). Initially, we observe the abnormal responses; as we take successive readings, they gradually decrease and move toward normal. After ongoing stimulation (T), the later responses appear to have returned to normal. Another record, obtained with platinum, displays the same pattern (fig. 77).


Fig. 76 and Fig. 77
Fig. 76 and Fig. 77
Abnormal ‘down’ response in tin (fig. 76) and in platinum (fig. 77) transformed into normal ‘up’ response, after continuous stimulation, T.
Abnormal 'down' response in tin (fig. 76) and in platinum (fig. 77) changed to normal 'up' response after continuous stimulation, T.
On placing the three sets of records—nerve, tin, and platinum—side by side, it will be seen how essentially similar they are in every respect.[17]
On putting the three sets of records—nerve, tin, and platinum—next to each other, you'll see just how similar they are in every way.
This reversion to normal is seen to have appeared in a pronounced manner after rapidly continuous stimulation, in process of which the modified molecular condition must in some way have reverted to the normal.
This return to normal is clearly noticeable after continuous stimulation, during which the altered molecular state must have somehow reverted to its original condition.

Fig. 78.—The Gradual Transition from Abnormal To Normal Response in Platinum
Fig. 78.—The Gradual Transition from Abnormal to Normal Response in Platinum
The transition will be seen to have commenced at the third and ended at the seventh, counting from the left.
The transition will be recognized as starting at the third and ending at the seventh, counting from the left.
Being desirous to trace this change gradually taking place, I took a platinum wire cell giving modified responses, and obtained a series of records of effects of individual stimuli continued for a long time. In this series, the points of transition from modified response to normal will be clearly seen (fig. 78).
Being eager to track this gradual change, I used a platinum wire cell that provided altered responses and gathered a set of records showing the effects of individual stimuli over an extended period. In this series, you can clearly see the points where the response shifts from modified to normal (fig. 78).

Fig. 79.—The Normal Response a in Nerve Enhanced to b after Continuous Stimulation T (Waller)
Fig. 79.—The Normal Response a in Nerve Boosted to b after Ongoing Stimulation T (Waller)
The normal response in nerve is recorded ‘down.’
The typical response in a nerve is recorded as 'down.'

Fig. 80.—Enhanced Response in Platinum after Continuous Stimulation T
Fig. 80.—Improved Response in Platinum after Ongoing Stimulation T
Increased response after continuous stimulation.—We have seen that responses to uniform stimuli sometimes show a staircase increase, apparently owing to the gradual removal of molecular sluggishness. Possibly analogous to this is the increase of response in nerve after continuous stimulation or tetanisation, observed by Waller (fig. 79). Like the staircase effect, this contravenes the commonly accepted theory of the dissimilation of tissue by stimulus, and the consequent depression of response. It is suggested by Waller that this increase of response after tetanisation may be due to the hypothetical evolution of CO2 to which allusion has previously been made.
Increased response after continuous stimulation.—We have observed that responses to consistent stimuli sometimes exhibit a staircase-like increase, likely due to the gradual reduction of molecular sluggishness. A similar increase in response has been noted in nerves after continuous stimulation or tetanization, as observed by Waller (fig. 79). Like the staircase effect, this challenges the widely accepted theory that tissue deteriorates due to stimulus, leading to a decrease in response. Waller suggests that this increase in response after tetanization might be because of the hypothetical release of CO2 that has been mentioned earlier.

Fig. 81.—Enhanced Response in Tin After Continuous Stimulation T
Fig. 81.—Improved Reaction in Tin After Ongoing Stimulation T
But there is an exact correspondence between this phenomenon and that exhibited by metals under similar conditions. I give here two sets of records (figs. 80, 81), one obtained with platinum and the other with tin, which demonstrate how the response is enhanced after continuous stimulation in a manner exactly similar to that noticed in the case of nerve.
But there is a direct connection between this phenomenon and what happens with metals under similar conditions. I'm providing two sets of records (figs. 80, 81), one collected with platinum and the other with tin, which show how the response improves after continuous stimulation in a way that mirrors what is observed in nerves.
The explanation which has been suggested with regard to the staircase effect—increased molecular mobility due to removal of sluggishness by repeated stimulation—would appear to be applicable in this case also. It would appear, then, that in all the phenomena which we have studied under the heads of ‘staircase’ effect, increase of response after continuous stimulation, and fatigue, there is a similarity between the observations made upon the response of muscle and nerve on the one hand, and that of metals on the other. Even in their abnormalities we have seen an agreement.
The explanation suggested regarding the staircase effect—increased molecular movement due to the removal of sluggishness through repeated stimulation—seems to apply here as well . It seems that in all the phenomena we've studied under the headings of 'staircase' effect, increased response after continuous stimulation, and fatigue, there is a similarity between the observations of muscle and nerve responses on one hand and those of metals on the other. Even in their abnormalities, we have observed a connection.
But amongst these phenomena themselves, though at first sight so diverse, there is some kind of continuity. Calling all normal response positive, for the sake of convenience, we observe its gradual modification, corresponding to changes in the molecular condition of the substance.
But among these phenomena, even though they seem so different at first, there is some kind of continuity. If we call all normal responses positive for convenience, we can see their gradual change in relation to the changes in the molecular state of the substance.
Beginning with that case in which molecular modification is extreme, we find a maximum variation of response from the normal, that is to say, to negative.
Beginning with that case where molecular modification is extreme, we see a maximum change in response from the normal, meaning to negative.
Continued stimulation, however, brings back the molecular condition to normal, as evidenced by the progressive lessening of the negative response, culminating in reversion to the normal positive. This is equally true of nerve and metal.
Continued stimulation, however, restores the molecular condition to normal, as shown by the gradual reduction of the negative response, eventually returning to the normal positive. This is also true for both nerves and metals.
In the next class of phenomena, the modification of molecular condition is not so great. It now exhibits itself merely as a relative inertness, and the responses, though positive, are feeble. Under continued stimulation, they increase in the same direction as in the last case, that is to say, from less positive to more positive, being the reverse of fatigue. This is evidenced alike by the staircase effect and by the increase of response after tetanisation, seen not only in nerve but also in platinum and tin.
In the next set of phenomena, the change in molecular condition isn't as significant. It now shows up mainly as a kind of relative inactivity, and the responses, while positive, are weak. With ongoing stimulation, they increase in the same way as before, moving from less positive to more positive, which is the opposite of fatigue. This is demonstrated by the staircase effect and by the increased response after tetanization, observed not just in nerves but also in platinum and tin.
The substance may next be in what we call the normal condition. Successive uniform stimuli now evoke uniform and equal positive responses, that is to say, there is no fatigue. But after intense or long-continued stimulation, the substance is overstrained. The responses now undergo a change from positive to less positive; fatigue, that is to say, appears.
The substance can next be in what we consider the normal state. Consistent and uniform stimuli now trigger equal and positive responses, meaning there is no fatigue. However, after intense or prolonged stimulation, the substance becomes overworked. The responses then shift from positive to less positive; in other words, fatigue sets in.
Again, under very much prolonged stimulation the response may decline to zero, or even undergo a reversal to negative, a phenomenon which we shall find instanced in the reversed response of retina under the long-continued stimulus of light.
Again, with prolonged stimulation, the response may drop to zero or even reverse to negative. This phenomenon can be seen in the reversed response of the retina after a long exposure to light.
We must then recognise that a substance may exist in various molecular conditions, whether due to internal changes or to the action of stimulus. The responses give us indications of these conditions. A complete cycle of molecular modifications can be traced, from the abnormal negative to the normal positive, and then again to negative seen in reversal under continuous stimulation.
We need to acknowledge that a substance can exist in different molecular states, whether because of internal changes or external stimuli. The responses signal these states to us. We can follow a complete cycle of molecular changes, moving from an abnormal negative state to a normal positive one, and then back to negative when there is ongoing stimulation.
FOOTNOTES:
[16] ‘Considering that we have no previous evidence of any chemical or physical change in tetanised nerve, it seems to me not worth while pausing to deal with the criticism that it is not CO2, but “something else” that has given the result.’—Waller, Animal Electricity, p. 59. That this phenomenon is nevertheless capable of physical explanation will be shown presently.
[16] "Since we have no prior evidence of any chemical or physical change in tetanized nerve, I don't think it's worth our time to respond to the criticism that it's not CO2, but rather 'something else' that caused the result." —Waller, Animal Electricity, p. 59. However, this phenomenon can still be explained physically, as will be shown shortly.
[17] In order to explain the phenomena of electric response, some physiologists assume that the negative response is due to a process of dissimilation, or breakdown, and the positive to a process of assimilation, or building up, of the tissue. The modified or positive response in nerve is thus held to be due to assimilation; after continuous stimulation, this process is supposed to be transformed into one of dissimilation, with the attendant negative response.
[17] To explain how the electric response works, some physiologists suggest that the negative response comes from a process of dissimilation, or breaking down, while the positive response is from a process of assimilation, or building up, of the tissue. The modified or positive response in nerves is thought to result from assimilation; after continuous stimulation, this process is believed to shift into dissimilation, leading to the negative response.
How arbitrary and unnecessary such assumptions are will become evident, when the abnormal and normal responses, and their transformation from one to the other, are found repeated in all details in metals, where there can be no question of the processes of assimilation or dissimilation.
How arbitrary and unnecessary these assumptions are will become clear when we see that abnormal and normal responses, along with their transformation from one to the other, are consistently observed in all aspects of metals, where there's no question of the processes of assimilation or dissimilation.
CHAPTER XV
INORGANIC RESPONSE—RELATION BETWEEN STIMULUS AND
RESPONSE—SUPERPOSITION OF STIMULI
- Relation between stimulus and response
- —Magnetic analogue
- —Increase of response with increasing Stimulus
- —Threshold of response
- —Superposition of Stimuli
- —Hysteresis.
Relation between stimulus and response.—We have seen what extremely uniform responses are given by tin, when the intensity of stimulus is maintained constant. Hence it is obvious that these phenomena are not accidental, but governed by definite laws. This fact becomes still more evident when we discover how invariably response is increased by increasing the intensity of stimulus.
Relation between stimulus and response.—We have observed how consistently tin responds when the intensity of the stimulus remains constant. Therefore, it is clear that these phenomena are not random but are controlled by specific laws. This fact becomes even more apparent when we find that increasing the intensity of the stimulus consistently leads to a stronger response.
Electrical response is due, as we have seen, to a molecular disturbance, the stimulus causing a distortion from a position of equilibrium. In dealing with the subject of the relation between the disturbing force and the molecular effect it produces, it may be instructive to consider certain analogous physical phenomena in which molecular deflections are also produced by a distorting force.
Electrical response occurs because, as we’ve observed, there is a molecular disturbance, with the stimulus leading to a shift from a state of balance. When examining the connection between the disturbing force and the molecular effects it creates, it might be helpful to look at certain similar physical phenomena where molecular shifts are also caused by a distorting force.
Magnetic analogue.—Let us consider the effect that a magnetising force produces on a bar of soft iron. It is known that each molecule in such a bar is an individual magnet. The bar as a whole, nevertheless, exhibits no external magnetisation. This is held to be due to the fact that the molecular magnets are turned either in haphazard directions or in closed chains, and there is therefore no resultant polarity. But when the bar is subjected to a magnetising force by means, say, of a solenoid carrying electrical current, the individual molecules are elastically deflected, so that all the molecular magnets tend to place themselves along the lines of magnetising force. All the north poles thus point more or less one way, and the south poles the other. The stronger the magnetising force, the nearer do the molecules approach to a perfect alignment, and the greater is the induced magnetisation of the bar.
Magnetic Analogue.—Let’s look at how a magnetizing force affects a bar of soft iron. It’s known that each molecule in such a bar acts like an individual magnet. However, the bar itself shows no external magnetization. This is thought to be because the molecular magnets are oriented in random directions or in closed loops, resulting in no overall polarity. But when the bar is exposed to a magnetizing force, like that from a solenoid with an electrical current, the individual molecules get elastically displaced, so all the molecular magnets try to align themselves with the lines of magnetizing force. All the north poles then point roughly in one direction, while the south poles point the other way. The stronger the magnetizing force, the closer the molecules get to perfect alignment, and the greater the induced magnetization of the bar.
The intensity of this induced magnetisation may be measured by noting the deflection it produces on a freely suspended magnet in a magnetometer.
The strength of this induced magnetization can be measured by observing the deflection it causes on a freely suspended magnet in a magnetometer.
The force which produces that molecular deflection, to which the magnetisation of the bar is immediately due, is the magnetising current flowing round the solenoid. The magnetisation, or the molecular effect, is measured by the deflection of the magnetometer. We may express the relation between cause and effect by a curve in which the abscissa represents the magnetising current, and the ordinate the magnetisation produced (fig. 82).
The force that causes the molecular deflection, which directly results in the magnetization of the bar, is the magnetizing current flowing through the solenoid. The magnetization, or the molecular effect, is measured by the deflection of the magnetometer. We can represent the relationship between cause and effect with a graph where the horizontal axis shows the magnetizing current, and the vertical axis shows the resulting magnetization (fig. 82).

In such a curve we may roughly distinguish three parts. In the first, where the force is feeble, the mole cular deflection is slight. In the next, the curve is rapidly ascending, i.e. a small variation of impressed force produces a relatively large molecular effect. And lastly, a limit is reached, as seen in the third part, where increasing force produces very little further effect. In this cause-and-effect curve, the first part is slightly convex to the abscissa, the second straight and ascending, and the third concave.
In this curve, we can basically see three parts. In the first part, where the force is weak, the molecular deflection is minimal. In the next part, the curve climbs quickly, meaning that a small change in applied force leads to a relatively large molecular response. Finally, a limit is hit in the third part, where increasing the force results in very little additional effect. In this cause-and-effect curve, the first part is slightly curved upward away from the horizontal axis, the second part is straight and rising, and the third part curves downward.
Increase of response with increasing stimulus.—We shall find in dealing with the relation between the stimulus and the molecular effect—i.e. the response—something very similar.
Increase of response with increasing stimulus.—We’ll discover that when looking at the relationship between the stimulus and the molecular effect—meaning the response—there's something quite similar happening.
On gradually increasing the intensity of stimulus, which may be done, as already stated, by increasing the amplitude of vibration, it will be found that, beginning with feeble stimulation, this increase is at first slight, then more pronounced, and lastly shows a tendency to approach a limit. In all this we have a perfect parallel to corresponding phenomena in animal and vegetable response. We saw that the proper investigation of this subject was much complicated, in the case of animal and vegetable tissues, by the appearance of fatigue. The comparatively indefatigable nature of tin causes it to offer great advantages in the pursuit of this inquiry. I give below two series of records made with tin. The first record, fig. 83, is for increasing amplitudes from 5° to 40° by steps of 5°. The stimuli are imparted at intervals of one minute. It will be noticed that whereas the recovery is complete in one minute when the stimulus is moderate, it is not quite complete when the stimulus is stronger. The recovery from the effect of stronger stimulus is more prolonged. Owing to want of complete recovery, the base line is tilted slightly upward. This slight displacement of the zero line does not materially affect the result, provided the shifting is slight.
As the intensity of the stimulus gradually increases, which can be done by raising the amplitude of the vibration, it becomes clear that starting with weak stimulation, the increase is initially small, then becomes more noticeable, and eventually approaches a limit. This pattern closely mirrors similar reactions seen in both animal and plant responses. We observed that investigating this subject is much more complex in the case of animal and plant tissues due to the factor of fatigue. The relatively fatigue-resistant nature of tin allows it to provide significant advantages in this investigation. Below are two series of records made with tin. The first record, fig. 83, shows increasing amplitudes from 5° to 40° in increments of 5°. The stimuli are applied at one-minute intervals. It can be seen that while recovery is complete in one minute for moderate stimuli, it is not quite complete for stronger stimuli. The recovery from stronger stimuli takes longer. Due to incomplete recovery, the baseline is slightly tilted upwards. This slight shift of the zero line does not significantly impact the result, as long as the change is minimal.

Fig. 83.—Records of Responses in Tin with Increasing Stimuli, Amplitudes of Vibration from 5° to 40°
Fig. 83.—Records of Responses in Tin with Increasing Stimuli, Amplitudes of Vibration from 5° to 40°
The vertical line to the right represents ·1 volt.
The vertical line on the right represents 1 volt.
Table showing the Increasing Electric Response due to
Increasing Amplitude of Vibration
| Vibration amplitude | E.M. variation |
|---|---|
| 5° | ·024 volt |
| 10° | ·057 " |
| 20° | ·111 " |
| 25° | ·143 " |
| 30° | ·170 " |
| 35° | ·187 " |
| 40° | ·204 " |

Fig. 84.—A Second Set of Records with a Different Specimen of Tin
Fig. 84.—A Second Set of Records with a Different Sample of Tin
The amplitudes of vibration are increased by steps of 10°, from 20° to 160°. (The deflections are reduced by interposing a high external resistance.)
The vibration amplitudes increase in increments of 10°, from 20° to 160°. (The deflections decrease by adding a high external resistance.)
The next figure (fig. 84) gives record of responses through a wider range. For accurate quantitative measurements it is preferable to wait till the recovery is complete. We may accomplish this within the limited space of the recording photographic plate by making the record for one minute; during the rest of recovery, the clockwork moving the plate is stopped and the galvanometer spot of light is cut off. Thus the next record starts from a point of completed recovery, which will be noticed as a bright spot at the beginning of each curve. With stimulation of high intensity, a tendency will be noticed for the responses to approach a limit.
The next figure (fig. 84) shows the responses over a wider range. For accurate quantitative measurements, it's better to wait until recovery is fully complete. We can achieve this within the limited space of the recording photographic plate by making the record for one minute; during the rest of the recovery, the mechanism that moves the plate is stopped and the galvanometer light is turned off. This way, the next record starts from a point of full recovery, which will be seen as a bright spot at the beginning of each curve. With high-intensity stimulation, there will be a trend for the responses to reach a limit.

Fig. 85.—Effect of Superposition on Tin
Fig. 85.—Impact of Superposition on Tin
A single stimulus produces the feeble effect shown in the first response. Superposition of 5, 9, 13 such stimuli produce the succeeding stronger responses.
A single stimulus creates the weak effect shown in the first response. Stacking 5, 9, or 13 of these stimuli results in the subsequent stronger responses.
Threshold of response.—There is a minimum intensity of stimulus below which there is hardly any visible response. We may regard this point as the threshold of response. Though apparently ineffective, the subliminal stimuli produce some latent effect, which may be demonstrated by their additive action. The record in fig. 85 shows how individually feeble stimuli become markedly effective by superposition.
Threshold of response.—There’s a minimum level of stimulus intensity below which there’s hardly any visible reaction. We can consider this point as the threshold of response. Even though they seem ineffective, subliminal stimuli create some hidden impact, which can be shown through their combined effect. The record in fig. 85 shows how individually weak stimuli can become significantly effective when layered together.
Superposition of stimuli.—The additive effect of succeeding stimuli will be seen from the above. The fusion of effect will be incomplete if the frequency of stimulation be not sufficiently great; but it will tend to be more complete with higher frequency of stimulation (fig. 86). We have here a parallel case to the complete and incomplete tetanus of muscles, under similar conditions.
Superposition of stimuli.—The combined effect of consecutive stimuli can be seen from the above. The blending of effects will be incomplete if the rate of stimulation isn't high enough; however, it will become more complete with a higher rate of stimulation (fig. 86). This parallels the complete and incomplete tetanus of muscles under similar conditions.
By the addition of these rapidly succeeding stimuli, a maximum effect is produced, and further stimulation adds nothing to this. The effect is balanced by a force of restitution. The response-curve thus rises to its maximum, after which the deflection is held as it were rigid, so long as the vibration is kept up.
By adding these quickly following stimuli, we achieve the maximum effect, and any additional stimulation doesn't contribute anything further. The effect is countered by a restorative force that brings it back. The response curve then reaches its peak, and as long as the vibration continues, the deflection remains almost fixed.
It was found that increasing intensities of single stimuli produced correspondingly increased responses. The same is true also of groups of stimuli. The maximum effect produced by superposition of stimuli increases with the intensity of the constituent stimuli.
It was found that stronger single stimuli led to stronger responses. The same is also true for groups of stimuli. The maximum effect from combining stimuli increases with the intensity of the individual stimuli.

Fig. 86.—Incomplete and Complete Fusion of Effect in Tin
Fig. 86.—Partial and Full Fusion of Effect in Tin
As the frequency of stimulation is increased the fusion becomes more and more complete. Vertical line to the right represents ·1 volt.
As the stimulation frequency goes up, the fusion becomes increasingly complete. The vertical line on the right represents ·1 volt.
Hysteresis.—Allusion has already been made to the increased responsiveness conferred by preliminary stimulation (see p. 127). Being desirous of finding out in what manner this is brought about, I took a series of observations for an entire cycle, that is to say, a series of observations were taken for maximum effects, starting from amplitude of vibration of 10° and ending in 100°, and backwards from 100° to 10°. Effect of hysteresis is very clearly seen (see A, fig. 87); there is a considerable divergence between the forward and return curves, the return curve being higher. On repeating the cycle several times, the divergence is found very much reduced, the wire on the whole is found to assume a more constant sensitiveness. In this steady condition, generally speaking, the sensitiveness for smaller amplitude of vibration is found to be greater than at the very beginning, but the reverse is the case for stronger intensity of stimulation.
Hysteresis.—We've already mentioned the increased responsiveness that comes from initial stimulation (see p. 127). Wanting to understand how this happens, I conducted a series of observations over a full cycle, meaning I took observations for maximum effects, starting with a vibration amplitude of 10° and going up to 100°, then back down from 100° to 10°. The effect of hysteresis is very clear (see A, fig. 87); there is a noticeable difference between the forward and return curves, with the return curve being higher. When repeating the cycle several times, the difference is found to be much smaller, and the wire generally shows more consistent sensitivity. In this stable condition, the sensitivity for smaller vibrations is typically greater than at the very start, while the opposite is true for higher levels of stimulation.

Fig. 87.—Cyclic Curve for Maximum Effects showing Hysteresis
Fig. 87.—Cyclic Curve for Maximum Effects showing Hysteresis
Effect of annealing.—I repeated the experiment with the same wire, after pouring hot water into the cell and allowing it to cool to the old temperature. From the cyclic curve (B, fig. 87) it will be seen (1) that the sensitiveness has become very much enhanced; (2) that there is relatively less divergence between the forward and return curves. Even this divergence practically disappeared at the third cycle, when the forward and backward curves coincided (C, fig. 87). The above results show in what manner the excitability of the wire is enhanced by purely physical means.
Effect of annealing.—I repeated the experiment with the same wire after pouring hot water into the cell and letting it cool back to the initial temperature. From the cyclic curve (B, fig. 87), it can be seen (1) that the sensitivity has significantly increased; (2) that there is comparatively less divergence between the forward and return curves. This divergence almost disappeared during the third cycle when the forward and backward curves lined up (C, fig. 87). These results demonstrate how the excitability of the wire is enhanced by purely physical means.
It is very curious to notice that addition of Na2CO3 solution (see Chap. XV—Action of Stimulants) produces enhancement of responsive power similar to that produced by annealing; that is to say, not only is there a great increase of sensitiveness, but there is also a reduction of hysteresis.
It’s interesting to see that adding Na2CO3 solution (see Chap. XV—Action of Stimulants) enhances responsive ability in a way that’s similar to what happens during annealing. In other words, not only does it significantly increase sensitivity, but it also decreases hysteresis.
CHAPTER XVI
INORGANIC RESPONSE—EFFECT OF CHEMICAL REAGENT
- Action of chemical reagents
- —Action of stimulants on metals
- —Action of depressants on metals
- —Effect of ‘poisons’ on metals
- —Opposite effect of large and small doses.
We have seen that the ultimate criterion of the physiological character of electric response is held to be its abolition when the substance is subjected to those chemical reagents which act as poisons.
We have seen that the key measure of the physiological nature of electric response is considered to be its elimination when the substance is exposed to chemical agents that act as poisons.

Fig. 88.—Action of Poison in Abolishing Response in Nerve (Waller)
Fig. 88.—How Poison Affects Nerve Response (Waller)
Action of chemical reagents.—Of these reagents, some are universal in their action, amongst which strong solutions of acids and alkalis, and salts like mercuric chloride, may be cited. These act as powerful toxic agents, killing the living tissue, and causing electric response to disappear. (See fig. 88.) It must, however, be remembered that there are again specific poisons which may affect one kind of tissue and not others. Poisons in general may be regarded as extreme cases of depressants. As an example of those which produce moderate physiological depression, potassium bromide may be mentioned, and this also diminishes electric response. There are other chemical reagents, on the other hand, which produce the opposite effect of increasing the excitability and causing a corresponding exaltation of electric response.
Action of chemical reagents.—Some of these reagents work universally, including strong solutions of acids and bases, as well as salts like mercuric chloride. These are powerful toxins that kill living tissue and cause the electric response to disappear. (See fig. 88.) However, it's important to note that there are specific poisons that may affect certain types of tissue and not others. Poisons, in general, can be seen as extreme forms of depressants. For example, potassium bromide is a substance that causes moderate physiological depression and also reduces electric response. Conversely, there are other chemical reagents that have the opposite effect, increasing excitability and leading to an elevated electric response.
We shall now proceed to inquire whether the response of inorganic bodies is affected by chemical reagents, so that their excitability is exalted by some, and depressed or abolished by others. Should it prove to be so, the last test will have been fulfilled, and that parallelism which has been already demonstrated throughout a wide range of phenomena, between the electric response of animal tissues on the one hand, and that of plants and metals on the other, will be completely established.
We will now look into whether the reaction of inorganic substances is influenced by chemical agents, such that some enhance their excitability while others diminish or eliminate it. If this turns out to be the case, the final test will have been met, and the parallel that has already been shown across a broad range of phenomena—between the electric response of animal tissues and that of plants and metals—will be fully confirmed.
Action of stimulants on metals.—We shall first study the stimulating action of various chemical reagents. The method of procedure is to take a series of normal responses to uniform stimuli, the electrolyte being water. The chemical reagent whose effect is to be observed is now added in small quantity to the water in the cell, and a second series of responses taken, using the same stimulus as before. Generally speaking, the influence of the reagent is manifested in a short period, but there may be occasional instances where the effect takes some time to develop fully. We must remember that by the introduction of the chemical reagent some change may be produced in the internal resistance of the cell. The effect of this on the deflection is eliminated by interposing a very high external resistance (from one to five megohms) in comparison with which the internal resistance of the cell is negligible. The fact that the introduction of the reagent did not produce any variation in the total resistance of the circuit was demonstrated by taking two deflections, due to a definite fraction of a volt, before and after the introduction of the reagent. These deflections were found equal.
Action of stimulants on metals.—We will first examine how different chemical reagents stimulate. The approach involves taking a series of consistent responses to uniform stimuli, with water as the electrolyte. A small amount of the chemical reagent being tested is added to the water in the cell, and a second series of responses is measured using the same stimulus as before. Generally, the reagent's effect shows up quickly, but sometimes it may take longer to fully develop. It’s important to note that introducing the chemical reagent might change the internal resistance of the cell. To eliminate the impact of this on the deflection, we use a very high external resistance (between one and five megohms), making the internal resistance of the cell negligible. The fact that adding the reagent did not alter the total resistance of the circuit was confirmed by measuring two deflections caused by a specific fraction of a volt, before and after the reagent was introduced, which were found to be equal.


Action of depressants.—Certain other reagents, again, produce an opposite effect. That is to say, they diminish the intensity of response. The record given on the next page (fig. 91) shows the depressing action of 10 per cent. solution of KBr on tin.
Action of depressants.—Some other substances, on the other hand, have the opposite effect. In other words, they reduce the intensity of the response. The record provided on the next page (fig. 91) illustrates the depressing effect of a 10 percent solution of KBr on tin.

Fig. 91.—Depressing Effect of KBr (10 per Cent.) on the Response of Tin
Fig. 91.—Depressive Effect of KBr (10 Percent) on the Response of Tin
Effect of ‘poison.’—Living tissues are killed, and their electric responses are at the same time abolished by the action of poisons. It is very curious that various chemical reagents are similarly effective in killing the response of metals. I give below a record (fig. 92) to show how oxalic acid abolishes the response. The depressive effect of this reagent is so great that a strength of one part in 10,000 is often sufficient to produce complete abolition. Another notable point with reference to the action of this reagent is the persistence of after-effect. This will be clearly seen from an account of the following experiment. The two wires A and B, in the cell filled with water, were found to give equal responses. The wires were now lifted off, and one wire B was touched with dilute oxalic acid. All traces of acid were next removed by rubbing the wire with cloth under a stream of water. On replacing the wire in the cell, A gave the usual response, whereas that of B was found to be abolished. The depression produced is so great and passes in so deep that I have often failed to revive the response, even after rubbing the wire with emery paper, by which the molecular layer on the surface must have been removed.
Effect of ‘poison.’—Living tissues are destroyed, and their electric responses are simultaneously eliminated by the action of poisons. It’s interesting that various chemical agents are similarly effective in stopping the response of metals. Below, I provide a record (fig. 92) to show how oxalic acid eliminates the response. The suppressive effect of this agent is so significant that a concentration of one part in 10,000 is often enough to completely eliminate it. Another important aspect regarding the effect of this agent is the lasting after-effect. This will be clearly illustrated by the following experiment. The two wires A and B, in the cell filled with water, were found to give equal responses. The wires were then removed, and one wire B was touched with dilute oxalic acid. All traces of acid were then wiped off by rubbing the wire with cloth under a stream of water. When the wire was placed back in the cell, A produced the usual response, while that of B was found to be eliminated. The level of depression caused is so profound and deep that I have often been unable to restore the response, even after rubbing the wire with emery paper, which would have removed the molecular layer from the surface.

We have seen in the molecular model (fig. 62, d, e) how the attainment of maximum is delayed, the response diminished, and the recovery prolonged or arrested by increase of friction or reduction of molecular mobility.
We have seen in the molecular model (fig. 62, d, e) how reaching the maximum is delayed, the response is weaker, and the recovery is extended or stopped by increased friction or decreased molecular mobility.
It would appear as if the reagents which act as poisons produced some kind of molecular arrest. The following records seen to lend support to this view. If the oxalic acid is applied in large quantities, the abolition of response is complete. But on carefully adding just the proper amount I find that the first stimulus evokes a responsive electric twitch, which is less than the normal, and the period of recovery is very much prolonged from the normal one minute before, to five minutes after, the application of the reagent (fig. 93, a). In another record the arrest is more pronounced, i.e. there is now no recovery (fig. 93, b). Note also that the maximum is attained much later. Stimuli applied after the arrest produce no effect, as if the molecular mechanism became, as it were, clogged or locked up.
It seems that the reagents acting as poisons caused some sort of molecular halt. The following records support this idea. When oxalic acid is applied in large amounts, the loss of response is total. However, if just the right amount is carefully added, I find that the initial stimulus produces a slight electric twitch that is less than normal, and the recovery time is significantly extended from the usual one minute to five minutes after the reagent is applied (fig. 93, a). In another record, the halt is more evident, meaning there’s no recovery at all (fig. 93, b). It's also worth noting that the maximum response occurs much later. Stimuli applied after this halt have no effect, as if the molecular mechanism has become, in a sense, jammed or locked.
In connection with this it is interesting to note that the effect of veratrine poison on muscle is somewhat similar. This reagent not only diminishes the excitability, but causes a very great prolongation of the period of recovery.
In relation to this, it's interesting to note that the effect of veratrine poison on muscle is somewhat similar. This substance not only reduces excitability but also leads to a significant extension of the recovery period.

Fig. 93.—‘Molecular Arrest’ by the Action of ‘poison’
Fig. 93.—‘Molecular Arrest’ by the Action of ‘poison’
In each, curves to the left show the normal response, curve to the right shows the effect of poison. In (a) the arrest is evidenced by prolongation of period of recovery. In (b) there is no recovery.
In each case, the curves that go left represent the normal response, while the curves that go right indicate the effect of poison. In (a), the arrest is shown by a longer recovery period. In (b), there is no recovery.
(1) The effect of these reagents is not only to increase or diminish the height of the response-curve, but also to modify the time relations. By the action of some the latent period is diminished, others produce a prolongation of the period of recovery. Some curious effects produced by the change of time relations have been noticed in the account given of diphasic variation (see p. 113).
(1) The impact of these substances is not just to raise or lower the response curve, but also to change the timing relationships. Some reduce the latent period, while others extend the recovery period. Some interesting effects caused by altering these timing relationships have been observed in the description of diphasic variation (see p. 113).
(3) A certain time is required for the full development of the effect. With some reagents the full effect takes place almost instantaneously, while with others the effect takes place slowly. Again the effect may with time reach a maximum, after which there may be a slight decline.
(3) It takes some time for the full effect to develop. For some reagents, the full effect happens almost instantly, while for others, it occurs more slowly. Additionally, the effect may reach a maximum after a while, and then there might be a slight decline.

Fig. 94.—Opposite Effects of Small and Large Doses (Tin)
Fig. 94.—Opposite Effects of Small and Large Doses (Tin)
(a) is the normal response; (b) is the stimulating action of small dose of potash (3 parts in 1,000); (c) is the abolition of response with a stronger dose (3 parts in 100).
(a) is the typical reaction; (b) is the stimulating effect of a small amount of potash (3 parts in 1,000); (c) is the elimination of the reaction with a higher dose (3 parts in 100).
(4) The after-effects of the reagents may be transitory or persistent; that is to say, in some cases the removal of the reagent causes the responses to revert to the normal, while in others the effect persists even after the removal of all traces of the reagent.
(4) The after-effects of the reagents can be temporary or long-lasting; that is to say, in some cases, taking away the reagent makes the responses go back to normal, while in others, the effect continues even after all traces of the reagent have been removed.
Opposite effects of large and small doses.—There remains a very curious phenomenon, known not only to students of physiological response but also known in medical practice, namely that of the opposite effects produced by the same reagent when given in large or in small doses. Here, too, we have the same phenomena reproduced in an extraordinary manner in inorganic response. The same reagent which becomes a ‘poison’ in large quantities may act as a stimulant when applied in small doses. This is seen in record fig. 94, in which (a) gives the normal responses in water; KHO solution was now added so as to make the strength three parts in 1,000, and (b) shows the consequent enhancement of response. A further quantity of KHO was added so as to increase the strength to three parts in 100. This caused a complete abolition (c) of response.
Opposite effects of large and small doses.—There remains a very curious phenomenon, known not only to students of physiological response but also known in medical practice, namely the opposite effects produced by the same substance when given in large or small doses. Here, too, we see the same phenomena appearing in a remarkable way in inorganic responses. The same substance that can become a ‘poison’ in large amounts may act as a stimulant when used in small doses. This is illustrated in record fig. 94, in which (a) exhibits the normal responses in water; KHO solution was then added to make the strength three parts per 1,000, and (b) shows the resulting enhancement of response. A further amount of KHO was added to increase the strength to three parts per 100. This led to a complete loss (c) of response.
It will thus be seen that as in the case of animal tissues and of plants, so also in metals, the electrical responses are exalted by the action of stimulants, lowered by depressants, and completely abolished by certain other reagents. The parallelism will thus be found complete in every detail between the phenomena of response in the organic and the inorganic.
It can therefore be observed that, just like in animal tissues and plants, metals also show enhanced electrical responses when stimulants are applied, decreased responses when depressants are used, and responses that are completely eliminated by certain other substances. This comparison will be found to be entirely consistent in every detail between the reactions in organic and inorganic materials.
CHAPTER XVII
ON THE STIMULUS OF LIGHT AND RETINAL CURRENTS
- Visual impulse: (1) chemical theory; (2) electrical theory
- —Retinal currents
- —Normal response positive
- —Inorganic response under stimulus of light
- —Typical experiment on the electrical effect induced by light.
The effect of the stimulus of light on the retina is perceived in the brain as a visual sensation. The process by which the ether-wave disturbance causes this visual impulse is still very obscure. Two theories may be advanced in explanation.
The effect of light on the retina is experienced in the brain as a visual sensation. The way the disturbance of ether waves creates this visual impulse is still quite unclear. Two theories can be proposed to explain it.
(1) Chemical theory.—According to the first, or chemical, theory, it is supposed that certain visual substances in the retina are affected by light, and that vision originates from the metabolic changes produced in these visual substances. It is also supposed that the metabolic changes consist of two phases, the upward, constructive, or anabolic phase, and the downward, destructive, or katabolic phase. Various visual substances by their anabolic or katabolic changes are supposed to produce the variations of sensation of light and colour. This theory, as will be seen, is very complex, and there are certain obstacles in the way of its acceptance. It is, for instance, difficult to see how this very quick visual process could be due to a comparatively slow chemical action, consisting of the destructive breaking-down of the tissue, followed by its renovation. Some support was at first given to this chemical theory by the bleaching action of light on the visual purple present in the retina, but it has been found that the presence or absence of visual purple could not be essential to vision, and that its function, when present, is of only secondary importance. For it is well known that in the most sensitive portion of the human retina, the fovea centralis, the visual purple is wanting; it is also found to be completely absent from the retinæ of many animals possessing keen sight.
(1) Chemical theory.—According to the first, or chemical, theory, it is believed that certain visual substances in the retina react to light, and that vision results from the metabolic changes that occur in these visual substances. These metabolic changes are thought to involve two phases: the upward, constructive, or anabolic phase, and the downward, destructive, or katabolic phase. Various visual substances are believed to create the differences in the sensation of light and color through their anabolic or katabolic changes. This theory is quite complex, and there are some challenges to its acceptance. For example, it's hard to understand how this very rapid visual process could be caused by a relatively slow chemical action involving the destructive breakdown of tissue, followed by its renewal. Initially, some evidence supported this chemical theory due to the bleaching effect of light on the visual purple found in the retina, but it has been determined that the presence or absence of visual purple is not crucial to vision, and that its role, when present, is only of secondary importance. It is well known that in the most sensitive part of the human retina, the fovea centralis, visual purple is absent; it is also completely missing in the retinas of many animals that have sharp vision.
(2) Electrical theory.—The second, or electrical, theory supposes that the visual impulse is the concomitant of an electrical impulse; that an electrical current is generated in the retina under the incidence of light, and that this is transmitted to the brain by the optic nerve. There is much to be said in favour of this view, for it is an undoubted fact, that light gives rise to retinal currents, and that, conversely, an electrical current suitably applied causes the sensation of light.
(2) Electrical theory.—The second, or electrical, theory suggests that the visual impulse happens alongside an electrical impulse; that an electrical current is created in the retina when light hits it, and this is sent to the brain through the optic nerve. There's a lot of evidence supporting this idea, as it's a well-established fact that light generates retinal currents, and, on the flip side, an electrical current applied correctly can produce the sensation of light.
Retinal currents.—Holmgren, Dewar, McKendrick, Kuhne, Steiner, and others have shown that illumination produces electric variation in a freshly excised eye. About this general fact of the electrical response there is a widespread agreement, but there is some difference of opinion as regards the sign of this response immediately on the application, cessation, and during the continuance of light. These slight discrepancies may be partly due to the unsatisfactory nomenclature—as regards use of terms positive and negative—hitherto in vogue and partly also to the differing states of the excised eyes observed.
Retinal currents.—Holmgren, Dewar, McKendrick, Kuhne, Steiner, and others have demonstrated that exposure to light creates an electrical change in a freshly removed eye. There's a general consensus about this electrical response, but opinions differ on the nature of this response immediately after light is applied, when it stops, and during its duration. These minor discrepancies may be partly due to unclear terminology concerning the use of the terms positive and negative that has been common in the past, and also to the varying conditions of the excised eyes being studied.
Waller, in his excellent and detailed work on the retinal currents of the frog, has shown how the sign of response is reversed in the moribund condition of the eye.
Waller, in his excellent and detailed study on the retinal currents of the frog, has demonstrated how the sign of response changes in the dying state of the eye.
As to the confusion arising from our present terminology, we must remember that the term positive or negative is used with regard to a current of reference—the so-called current of injury.
As for the confusion coming from our current terminology, we need to keep in mind that the terms positive and negative are used with respect to a reference current—the so-called current of injury.

Fig. 95. Retinal Response To Light
Fig. 95. Retinal Response to Light
The current of response is from the nerve to the retina.
The flow of response goes from the nerve to the retina.
When the two galvanometric contacts are made, one with the cut end of the nerve, and the other on the uninjured cornea, a current of injury is found which in the eye is from the nerve to the retina. In the normal freshly excised eye, the current of response due to the action of light on the retina is always from the nerve, which is not directly stimulated by light, to the retina, that is, from the less excited to the more excited (fig. 95). This current of response flows, then, in the same direction as the existing current of reference—the current of injury—and may therefore be called positive. Unfortunately the current of injury is very often apt to change its sign; it then flows through the eye from the cornea to the nerve. And now, though the current of response due to light may remain unchanged in direction, still, owing to the reversal of the current of reference, it will appear as negative. That is to say, though its absolute direction is the same as before, its relative direction is altered.
When the two galvanometric contacts are established, one on the cut end of the nerve and the other on the uninjured cornea, a current of injury is detected that in the eye flows from the nerve to the retina. In a normal freshly excised eye, the current of response caused by light acting on the retina always moves from the nerve, which isn't directly stimulated by light, to the retina, meaning from the less excited to the more excited (fig. 95). This response current, therefore, flows in the same direction as the existing reference current—the current of injury—and can be termed positive. Unfortunately, the current of injury often tends to change its sign; at that point, it flows through the eye from the cornea to the nerve. Now, even though the current of response due to light may remain unchanged in direction, due to the reversal of the reference current, it will appear as negative. In other words, while its absolute direction remains the same as before, its relative direction has shifted.
I have already advocated the use of the term positive for currents which flow towards the stimulated, and negative for those whose flow is away from the stimulated. If such a convention be adopted, no confusion can arise, even when, as in the given cases, the currents of injury undergo a change of direction.
Normal response positive.—The normal effect of light on the retina, as noticed by all the observers already mentioned, is a positive variation, during exposure to light of not too long duration. Cessation of light is followed by recovery. On these points there is general agreement amongst investigators. Deviations are regarded as due to abnormal conditions of the eye, owing to rough usage, or to the rapid approach of death. For just as in the dying plant we found occasional reversals from negative to positive response, so in the dying retina the response may undergo changes from the normal positive to negative.
Normal response positive.—The usual effect of light on the retina, as observed by all the previously mentioned observers, is a positive change during exposure to light for a short duration. When the light stops, recovery follows. There is a general consensus among researchers on these points. Any deviations are considered to result from abnormal conditions of the eye, caused by rough handling or nearing death. Just as we observed occasional shifts from negative to positive response in a dying plant, the dying retina may also experience changes from the normal positive response to negative.
The sign of response, as we have already seen in numerous cases, depends very much on the molecular condition of the sensitive substance, and if this condition be in any way changed, it is not surprising that the character of the response should also undergo alteration.
The sign of response, as we've already observed in many instances, heavily depends on the molecular state of the sensitive substance, and if this state changes in any way, it's no surprise that the nature of the response would also change.
Unlike muscle in this, successive retinal responses exhibit little change, for, generally speaking, fatigue is very slight, the retina recovering quickly even under strong light if the exposure be not too long. In exceptional cases, however, fatigue, or its converse, the staircase effect, may be observed.
Unlike muscle in this, successive retinal responses show little change, because, generally speaking, fatigue is minimal. The retina recovers quickly even under bright light as long as the exposure isn't too long. However, in rare cases, fatigue or its opposite, the staircase effect, can be observed.
Inorganic response under the stimulus of light.—It may now be asked whether such a complex vital phenomenon as retinal response could have its counterpart in non-living response. Taking a rod of silver, we may beat out one end into the form of a hollow cup, sensitising the inside by exposing it for a short time to vapour of bromine. The cup may now be filled with water, and connection made with a galvanometer by non-polarisable electrodes. There will now be a current due to difference between the inner surface and the rod. This may be balanced, however, by a compensating E.M.F.
Inorganic response to light stimuli.—We can now ask whether a complex vital phenomenon like the retinal response could have an equivalent in non-living systems. If we take a silver rod and shape one end into a hollow cup, sensitizing the interior by briefly exposing it to bromine vapor, we can then fill the cup with water and connect it to a galvanometer using non-polarizable electrodes. This will create a current resulting from the difference between the inner surface and the rod. However, this can be balanced by a compensating E.M.F.

Fig. 96.—Record of Responses To Light given by the Sensitive Cell
Fig. 96.—Record of Responses to Light given by the Sensitive Cell
Thick lines represent the effect during illumination, dotted lines the recovery in darkness. Note the preliminary negative twitch, which is sometimes also observed in responses of frog’s retina.
Thick lines show the effect during light exposure, while dotted lines indicate the recovery in the dark. Take note of the initial negative twitch, which is also sometimes seen in the responses of a frog's retina.
We have thus an arrangement somewhat resembling the eye, with a sensitive layer corresponding to the retina, and the less sensitive rod corresponding to the conducting nerve-stump (fig. 96, a).
We have an arrangement that looks a bit like the eye, with a sensitive layer that acts like the retina and the less sensitive rod resembling the nerve stump (fig. 96, a).
The apparatus is next placed inside a black box, with an aperture at the top. By means of an inclined mirror, light may be thrown down upon the sensitive surface through the opening.
The device is then put inside a black box, which has an opening at the top. Using a slanted mirror, light can be directed down onto the sensitive surface through the hole.
On exposing the sensitive surface to light, the balance is at once disturbed, and a responsive current of positive character produced. The current, that is to say, is from the less to the more stimulated sensitive layer. On the cessation of light, there is fairly quick recovery (fig. 96, b).
On exposing the sensitive surface to light, the balance is immediately disturbed, and a positive current is generated. This current flows from the less stimulated sensitive layer to the more stimulated one. When the light stops, recovery happens fairly quickly (fig. 96, b).
The character and the intensity of E.M. variation of the sensitive cell depend to some extent on the process of preparation. The particular cell with which most of the following experiments were carried out usually gave rise to a positive variation of about ·008 volt when acted on for one minute by the light of an incandescent gas-burner which was placed at a distance of 50 cm.
The character and intensity of the E.M. variation of the sensitive cell depend somewhat on how it is prepared. The specific cell used in most of the following experiments typically produced a positive variation of about 0.008 volts when exposed for one minute to the light of an incandescent gas burner positioned 50 cm away.

Fig. 97 (a)
Fig. 97 (a)
A, B are the two faces of a brominated sheet of silver. One face, say A, is acted on by light. The current of response is from B to A, across the plate.
A, B are the two sides of a brominated sheet of silver. One side, say A, is exposed to light. The flow of response is from B to A, across the plate.
Typical experiment on the electrical effect induced by light.—This subject of the production of an electrical current by the stimulus of light would appear at first sight very complex. But we shall be able to advance naturally to a clear understanding of its most complicated phenomena if we go through a preliminary consideration of an ideally simple case. We have seen, in our experiments on the mechanical stimulation of, for example, tin, that a difference of electric potential was induced between the more stimulated and less stimulated parts of the same rod, and that an action current could thus be obtained, on making suitable electrolytic connections. Whether the more excited was zincoid or cuproid depended on the substance and its molecular condition.
Typical experiment on the electrical effect induced by light.—At first glance, the idea of generating an electrical current using light may seem quite complex. However, we can gradually reach a clear understanding of its most intricate phenomena by first considering a straightforward case. In our experiments with the mechanical stimulation of materials like tin, we observed that a difference in electrical potential was created between the more stimulated and less stimulated areas of the same rod. This allowed us to obtain an action current when we made appropriate electrolytic connections. Whether the more excited area was zincoid or cuproid depended on the material and its molecular state.
Let us now imagine the metal rod flattened into a plate, and one face stimulated by light, while the other is protected. Would there be a difference of potential induced between the two faces of this same sheet of metal?
Let’s now picture the metal rod flattened into a plate, with one side exposed to light while the other side is shielded. Would this create a difference in voltage between the two sides of the plate?
Let two blocks of paraffin be taken and a large hole drilled through both. Next, place a sheet of metal between the blocks, and pour melted paraffin round the edge to seal up the junction, the two open ends being also closed by panes of glass. We shall have then two compartments separated by the sheet of metal, and these compartments may be filled with water through the small apertures at the top (fig. 97, a).
Take two blocks of paraffin and drill a large hole through both. Then, place a sheet of metal between the blocks and pour melted paraffin around the edge to seal the connection, with the two open ends also closed off by glass panes. This will create two compartments separated by the metal sheet, and these compartments can be filled with water through the small openings at the top (fig. 97, a).
Fig. 97 (b).—Record of Responses obtained from the Above Cell
Fig. 97 (b).—Record of Responses obtained from the Above Cell
Ten seconds’ exposure to light followed by fifty seconds’ recovery in the dark. Thick lines represent action in light, dotted lines represent recovery.
Ten seconds of light followed by fifty seconds of recovery in the dark. Solid lines show activity in light, while dotted lines indicate recovery.
The two liquid masses in the separated chambers thus make perfect electrolytic contacts with the two faces A and B of the sheet of metal. These two faces may be put in connection with a galvanometer by means of two non-polarisable electrodes, whose ends dip into the two chambers. If the sheet of metal have been properly annealed, there will now be no difference of potential between the two faces, and no current in the galvanometer. If the two faces are not molecularly similar, however, there will be a current, and the electrical effects to be subsequently described will act additively, in an algebraical sense. Let one face now be exposed to the stimulus of light. A responsive current will be found to flow, from the less to the more stimulated face, in some cases, and in others in an opposite direction.
The two liquid masses in the separate chambers create perfect electrolytic contacts with the two surfaces A and B of the metal sheet. These surfaces can be connected to a galvanometer using two non-polarisable electrodes, which are submerged in the two chambers. If the metal sheet has been properly annealed, there will be no difference in potential between the two surfaces, resulting in no current in the galvanometer. However, if the two surfaces are not molecularly identical, there will be a current, and the electrical effects described later will combine algebraically. Now, if one surface is exposed to light, a responsive current will be observed flowing from the less stimulated surface to the more stimulated one in some cases, while in others, it will flow in the opposite direction.
It appears at first very curious that this difference of electric potential should be maintained between opposite faces of a very thin and highly conducting sheet of metal, the intervening distance between the opposed surfaces being so extremely small, and the electrical resistance quite infinitesimal. A homogeneous sheet of metal has become by the unequal action of light, molecularly speaking, heterogeneous. The two opposed surfaces are thrown into opposite kinds of electric condition, the result of which is as if a certain thickness of the sheet, electrically speaking, were made zinc-like, and the rest copper-like. From such unfamiliar conceptions, we shall now pass easily to others to which we are more accustomed. Instead of two opposed surfaces, we may obtain a similar response by unequally lighting different portions of the same surface. Taking a sheet of metal, we may expose one half, say A, to light, the other half, B, being screened. Electrolytic contacts are made by plunging the two limbs in two vessels which are in connection with the two non-polarisable electrodes E and E′ (fig. 98, a). On illumination of A and B alternately, we shall now obtain currents flowing alternately in opposite directions.
At first, it seems quite strange that there should be a difference in electric potential between opposite sides of a very thin and highly conductive metal sheet, especially since the distance between the two surfaces is extremely small and the electrical resistance is nearly nonexistent. A uniform metal sheet has become, at a molecular level, uneven due to the unequal effect of light. The two opposite surfaces develop opposite types of electric states, resulting in a scenario where part of the sheet behaves like zinc and the rest behaves like copper. From these unfamiliar ideas, we can easily move to concepts that are more familiar to us. Instead of having two opposite surfaces, we can achieve a similar effect by unevenly lighting different areas of the same surface. If we take a metal sheet and expose one half, say A, to light while shielding the other half, B, we can create electrolytic contacts by immersing the two ends in two containers connected to the two non-polarizable electrodes E and E′ (fig. 98, a). By illuminating A and B alternately, we can then generate currents that flow alternately in opposite directions.

Just as in the strain cells the galvanometer contact was transferred from the electrolytic part to the metallic part of the circuit, so we may next, in an exactly similar manner, cut this plate into two, and connect these directly to the galvanometer, electrolytic connection being made by partially plunging them into a cell containing water. The posterior surfaces of the two half-plates may be covered with a non-conducting coating. And we arrive at a typical photo-electric cell (fig. 98, b). These considerations will show that the eye is practically a photo-electric cell.
Just like in the strain cells where the galvanometer contact was moved from the electrolytic part to the metallic part of the circuit, we can similarly split this plate in half and connect these directly to the galvanometer, making the electrolytic connection by partially immersing them in a cell filled with water. The back surfaces of the two half-plates can be covered with a non-conductive layer. This leads us to a typical photoelectric cell (fig. 98, b). These points demonstrate that the eye essentially functions as a photoelectric cell.

Fig. 99.—Responses To Light in Frog’s Retina
Fig. 99.—Responses to Light in Frog's Retina
Illumination L for one minute, recovery in dark for two minutes during obscurity D. (Waller.)
Illumination L for one minute, recovery in the dark for two minutes during obscurity D. (Waller.)
We shall now give detailed experimental results obtained with the sensitive silver-bromide cell, and compare its response-curve with those of the retina. A series of uniform light stimuli gives rise to uniform responses, which show very little sign of fatigue. How similar these response-curves are to those of the retina will be seen from a pair of records given below, where fig. 99 shows responses of frog’s retina, and fig. 100 gives the responses obtained with the sensitive silver cell (fig. 100).
We will now present detailed experimental results obtained from the sensitive silver-bromide cell and compare its response curve with those of the retina. A series of uniform light stimuli produces consistent responses, which show minimal signs of fatigue. The similarity of these response curves to those of the retina will be evident from the pair of records shown below, where fig. 99 shows responses from the frog’s retina, and fig. 100 displays the responses obtained from the sensitive silver cell (fig. 100).
It was said that the responses of the retina are uniform. This is only approximately true. In addition to numerous cases of uniform responses, Waller finds instances of ‘staircase’ increase, and its opposite, slight fatigue. In the record here given of the silver cell, the staircase effect is seen at the beginning, and followed by slight fatigue. I have other records where for a very long time the responses are perfectly uniform, there being no sign of fatigue.
It was said that the responses of the retina are consistent. This is only somewhat true. Along with many cases of consistent responses, Waller finds examples of a 'staircase' increase and its opposite, slight fatigue. In the record shown here of the silver cell, the staircase effect is observed at the beginning, followed by slight fatigue. I have other records where, for a very long time, the responses are perfectly consistent, showing no signs of fatigue.

Fig. 100.—Responses in Sensitive Silver Cell
Fig. 100.—Reactions in Sensitive Silver Cell
Illumination for one minute and obscurity for one minute. Thick line represents record during illumination, dotted line recovery during obscurity.
Illumination for one minute and darkness for one minute. The solid line shows the record during illumination, while the dotted line indicates recovery during darkness.
Another curious phenomenon sometimes observed in the response of retina is an occasional slight increase of response immediately on the cessation of light, after which there is the final recovery. An indication of this is seen in the second and fourth curves in fig. 99. Curiously enough, this abnormality is also occasionally met with in the responses of the silver cell, as seen in the first two curves of fig. 100. Other instances will be given later.
Another interesting phenomenon that can sometimes be seen in the retina's response is a slight increase in response right after the light stops, followed by a final recovery. This is shown in the second and fourth curves in fig. 99. Interestingly, this abnormality is also sometimes found in the responses of the silver cell, as shown in the first two curves of fig. 100. More examples will be provided later.
CHAPTER XVIII
INORGANIC RESPONSE—INFLUENCE OF VARIOUS CONDITIONS ON THE RESPONSE TO
STIMULUS OF LIGHT
- Effect of temperature
- —Effect of increasing length of exposure
- —Relation between intensity of light and magnitude of response
- —After-oscillation
- —Abnormal effects: (1) preliminary negative twitch; (2) reversal of response; (3) transient positive twitch on cessation of light; (4) decline and reversal
- —Résumé.
We shall next proceed to study the effect, on the response of the sensitive cell, of all those conditions which influence the normal response of the retina. We shall then briefly inquire whether even the abnormalities sometimes met with in retinal responses have not their parallel in the responses given by the inorganic.
We will now examine how different conditions affect the response of the sensitive cell, particularly those that influence the normal response of the retina. After that, we will briefly explore whether the abnormalities sometimes observed in retinal responses also have parallels in the responses of inorganic substances.

Fig. 101.—Influence of Temperature on Response
Fig. 101.—Effect of Temperature on Response
Illumination 20″, obscurity 40″.
Light 20″, dark 40″.
In (a) is shown a series of responses at 20° C.—the record exhibits slight fatigue. (b) is the slight irregular response at 50° C. (c) is the record on re-cooling; it exhibits ‘staircase’ increase.
In (a) is shown a series of responses at 20° C.—the record shows slight fatigue. (b) is the slight irregular response at 50° C. (c) is the record on re-cooling; it shows a 'staircase' increase.
Effect of temperature.—It has been found that when the temperature is raised above a certain point, retinal response shows rapid diminution. On cooling, however, response reappears, with its original intensity. In the response given by the sensitive cell, the same peculiarity is noticed. I give below (fig. 101, a) a set of response-curves for 20° C. These responses, after showing slight fatigue, became fairly constant. On raising the temperature to 50° C. response practically disappeared (101, b). But on cooling to the first temperature again, it reappeared, with its original if not slightly greater intensity (fig. 101, c). A curious point is that while in record (a), before warming, slight fatigue is observed, in (c), after cooling, the reverse, or staircase effect, appears.
Effect of temperature.—It has been found that when the temperature is raised above a certain point, the retinal response quickly decreases. However, when cooled down, the response comes back with its original intensity. The same pattern is seen in the response given by the sensitive cell. Below, I provide a set of response curves for 20° C (fig. 101, a). These responses, after showing slight fatigue, became fairly stable. When the temperature was increased to 50° C, the response almost vanished (101, b). But when cooled back down to the original temperature, it reappeared with its original intensity, if not slightly greater (fig. 101, c). An interesting point is that while in record (a), before warming, slight fatigue is observed, in (c), after cooling, the opposite, or staircase effect, appears.

Fig. 102.—Response-curves for Increasing Duration of Illumination from 1″ to 10″
Fig. 102.—Response curves for increased duration of illumination from 1 second to 10 seconds
In (a) the source of light was at a distance of 50 cm.; in (b) it was at a distance of 25 cm. Note the after-oscillation.
In (a) the light source was 50 cm away; in (b) it was 25 cm away. Note the after-oscillation.
Effect of increasing length of exposure.—If the intensity of light be kept constant, the magnitude of response of the sensitive cell increases with length of exposure. But this soon reaches a limit, after which increase of duration does not increase magnitude of effect. Too long an exposure may however, owing to fatigue, produce an actual decline.
Effect of Increasing Length of Exposure.—If the intensity of light remains constant, the response of the sensitive cell increases with the length of exposure. However, this soon hits a limit, after which extending the duration does not enhance the effect. An exposure that is too long may, due to fatigue, actually cause a decline.
I give here two sets of curves (fig. 102) illustrating the effect of lengthening exposure. The intensities of light in the two cases are as 1 to 4. The incandescent burner was in the two cases at distances 50 and 25 cm. respectively. It will be observed that beyond eight seconds’ exposure the responses are approximately uniform. Another noticeable fact is that with long exposure there is an after-oscillation. This growing effect with lengthening exposure and attainment of limit is exactly paralleled by responses of retina under similar conditions.
I present two sets of curves (fig. 102) showing the impact of longer exposure times. The light intensities in both cases are in a ratio of 1 to 4. The incandescent bulb was positioned at distances of 50 cm and 25 cm, respectively. It's noticeable that after eight seconds of exposure, the responses become fairly uniform. Another important point is that with extended exposure, there is an after-oscillation. This increasing effect with longer exposure and reaching a limit directly mirrors the responses of the retina under similar conditions.
Relation between intensity of light and magnitude of response.—In the responses of retina, it is found that increasing intensity of light produces an increasing effect. But the rate of increase is not uniform: increase of effect does not keep pace with increase of stimulus. Thus a curve giving the relation between stimulus and response is concave to the axis which represents the stimulus.
Relationship between light intensity and response magnitude.—In retinal responses, it is observed that higher light intensity leads to a greater effect. However, the rate of increase isn't consistent: the increase in effect doesn't match the increase in stimulus. Therefore, a graph representing the relationship between stimulus and response is concave to the axis that represents the stimulus.
The same is true of the sensation of light. That is to say, within wide limits, intensity of sensation does not increase so rapidly as stimulus.
The same is true for the feeling of light. In other words, within a broad range, the intensity of sensation doesn’t increase as quickly as the stimulus.
This particular relation between stimulus and effect is also exhibited in a remarkable manner by the sensitive cell. For a constant source of light I used an incandescent burner, and graduated the intensity of the incident light by varying its distance from the sensitive cell. The intensity of light incident on the cell, when the incandescent burner is at a distance of 150 cm., has been taken as the arbitrary unit. In order to make allowance for the possible effects of fatigue I took two successive series of responses (fig. 103). In the first, records were taken with intensities diminishing from 7 to 1, and immediately afterwards increasing from 1 to 7, in the second.
This specific relationship between stimulus and effect is also clearly shown by the sensitive cell. For a constant light source, I used an incandescent bulb and adjusted the intensity of the light hitting the cell by changing its distance. The intensity of light hitting the cell when the incandescent bulb is 150 cm away has been considered the arbitrary unit. To account for potential fatigue effects, I conducted two consecutive series of responses (fig. 103). In the first series, I recorded intensities decreasing from 7 to 1, and then immediately afterward, I recorded intensities increasing from 1 to 7 in the second series.

Fig. 103.—Responses of Sensitive Cell to various Intensities of Light
Fig. 103.—Responses of Sensitive Cell to Different Light Intensities
On the left the responses are for diminishing intensities in the ratios of 7, 5, 3, and 1. On the right they are for the increasing intensities 1, 3, 5, and 7. The thick lines are records during exposures of one minute; the dotted lines represent recoveries for one minute.
On the left, the responses are for lower intensities in the ratios of 7, 5, 3, and 1. On the right, they are for higher intensities of 1, 3, 5, and 7. The solid lines show data recorded during one-minute exposures; the dotted lines indicate recoveries over one minute.
Table giving Response to varying Intensities of Light
(The intensity of an incandescent gas-burner at a distance of 150 cm.
is taken as unit.)
(The intensity of an incandescent gas burner at a distance of 150 cm.
is considered the standard unit.)
| Intensity of Light |
Response (Light diminishing) |
Response (Light increasing) |
Mean | Value in volt |
|---|---|---|---|---|
| 7 | 43 | 39 | 41 | 63·0 × 10− volt |
| 5 | 31 | 29 | 30 | 46·1 ×The text appears to be empty. Please provide a short piece of text for me to modernize. |
| 3 | 18·5 | 17·5 | 18 | 27·7 × |
| 1 | 10 | 9 | 9·5 | 14·6 ×" |
As the zero point was slightly shifted during the course of the experiment, the deflection in each curve was measured from a line joining the beginning of the response to the end of its recovery. A mean deflection, corresponding to each intensity, was obtained by taking the average of the descending and ascending readings. The two sets of readings did not, however, vary to any marked extent.
As the starting point was slightly adjusted during the course of the experiment, the deflection in each curve was measured from a line connecting the start of the response to the end of its recovery. A mean deflection, corresponding to each intensity, was calculated by averaging the descending and ascending readings. However, the two sets of readings did not vary significantly.
The deflections corresponding to the intensities 1, 3, 5, 7, are, then, as 9·5 to 18, to 30, to 41. If the deflections had been strictly proportionate to the intensities of light stimulus they would have been as 9·5 to 28·5, to 47·5, to 66·5.
The deflections for the intensities 1, 3, 5, and 7 are 9.5, 18, 30, and 41, respectively. If the deflections had been exactly proportional to the intensities of the light stimulus, they would have been 9.5, 28.5, 47.5, and 66.5.

Fig. 104.—Curves giving the Relation between Intensity of Light and Magnitude of Response
Fig. 104.—Graphs Showing the Relationship Between Light Intensity and Response Magnitude
In (a) sensitive cell, (b) in frog’s retina.
In (a) a sensitive cell, (b) in a frog's retina.
In another set of records, with a different cell, I obtained the deflections of 6, 10, 13, 15, corresponding to light intensities of 3, 5, 7, and 9.
In another set of records, with a different cell, I got the deflections of 6, 10, 13, 15, corresponding to light intensities of 3, 5, 7, and 9.
The two curves in fig. 104, giving the relation between response and stimulus, show that in the case of inorganic substances, as in the retina (Waller), magnitude of response does not increase so rapidly as stimulus.
The two curves in fig. 104, showing the relationship between response and stimulus, indicate that for inorganic substances, like in the retina (Waller), the response magnitude doesn't increase as quickly as the stimulus.
After-oscillation.—When the sensitive surface is subjected to the continued action of light, the E.M. effect attains a maximum at which it remains constant for some time. If the exposure be maintained after this for a longer period, there will be a decline, as we found to be the case in other instances of continued stimulation. The appearance of this decline, and its rapidity, depends on the particular condition of the substance.
After-oscillation.—When the sensitive surface is exposed to light for a while, the E.M. effect reaches a maximum and stays steady for a bit. If the exposure continues beyond this point for a longer time, the effect will start to drop off, just like we found in other cases of ongoing stimulation. How quickly this decline appears and how fast it happens depends on the specific condition of the material.
When the sensitive element is considerably strained by the action of light, and if that light be now cut off, there is a rebound towards recovery and a subsequent after-oscillation. That is to say, the curve of recovery falls below the zero point, and then slowly oscillates back to the position of equilibrium. We have already seen an instance of this in fig. 102. Above is given a series of records showing the appearance of decline, from too long-continued exposure and recovery, followed by after-oscillation on the cessation of light (fig. 105). Certain visual analogues to this phenomenon will be noticed later.
When the sensitive element is significantly strained by light, and that light is suddenly turned off, there is a rebound towards recovery and a following after-oscillation. This means the recovery curve dips below the zero point, and then gradually oscillates back to the equilibrium position. We’ve already seen an example of this in fig. 102. Above is a series of records showing the decline from prolonged exposure and recovery, followed by after-oscillation once the light stops (fig. 105). Certain visual parallels to this phenomenon will be discussed later.

Fig. 105.—After-oscillation
Fig. 105.—After-oscillation
Exposure of one minute followed by obscurity of one minute. Note the decline during illumination, and after-oscillation in darkness.
Exposure of one minute followed by one minute of darkness. Observe the drop during the light, and the aftereffects in the dark.
Abnormal effects.—We have already treated of all the normal effects of the stimulus of light on the retina, and their counterparts in the sensitive cell. But the retina undergoes molecular changes when injured, stale, or in a dying condition, and under these circumstances various complicated modifications are observed in the response.
Abnormal effects.—We have already discussed all the normal effects of light stimulation on the retina, as well as their counterparts in the sensitive cell. However, the retina undergoes molecular changes when it is damaged, outdated, or dying, and in these situations, we see various complex changes in its response.

Fig. 106.—Transient Positive Augmentation given by the Frog’s Retina on the Cessation of Light L (Waller)
Fig. 106.—Temporary Positive Increase caused by the Frog’s Retina when Light L is Turned Off (Waller)

Fig. 107.—Responses in Silver Cell
Fig. 107.—Responses in Silver Cell
The thick line represents response during light (half a minute’s exposure), and dotted line the recovery during darkness. Note the terminal positive twitch.
The solid line shows the response during light (30 seconds of exposure), and the dotted line indicates the recovery during darkness. Pay attention to the final positive twitch.
1. Preliminary negative twitch.—When the light is incident on the frog’s retina, there is sometimes a transitory negative variation, followed by the normal positive response. This is frequently observed in the sensitive cell (see fig. 96, b).
1. Preliminary negative twitch.—When light hits the frog's retina, there can be a brief negative change, followed by the usual positive response. This is commonly seen in the sensitive cell (see fig. 96, b).
2. Reversal of response.—Again, in a stale retina, owing to molecular modification the response is apt to undergo reversal (Waller). That is to say, it now becomes negative. In working with the same sensitive cell on different days I have found it occasionally exhibiting this reversed response.
2. Reversal of response.—Once more, in an aged retina, due to molecular changes, the response tends to reverse (Waller). This means it becomes negative. When working with the same sensitive cell on different days, I've sometimes noticed it showing this reversed response.
3. Transient rise of current on cessation of light.—Another very curious fact observed in the retina by Kuhne and Steiner is that immediately on the stoppage of light there is sometimes a sudden increase in the retinal current, before the usual recovery takes place. This is very well shown in the series of records taken by Waller (fig. 106). It will be noticed that on illumination the response-curve rises, that continued illumination produces a decline, and that on the cessation of light there is a transient rise of current. I give here a series of records which will show the remarkable similarity between the responses of the cell and retina, in respect even of abnormalities so marked as those described (fig. 107). I may mention here that some of these curious effects, that is to say, the preliminary negative twitch and sudden augmentation of the current on the cessation of light, have also been noticed by Minchin in photo-electric cells.
3. Temporary spike in current when light stops.—Another intriguing observation made by Kuhne and Steiner in the retina is that right after the light shuts off, there can be a sudden spike in the retinal current before it starts to recover as usual. This is clearly illustrated in the series of recordings by Waller (fig. 106). You'll see that when illuminated, the response-curve goes up, that continued light causes it to decline, and that when the light stops, there’s a temporary increase in current. I’m providing a series of records that highlight the striking similarities between the responses of the cell and retina, even regarding noticeable abnormalities like those discussed (fig. 107). It's worth noting that some of these odd effects, specifically the preliminary negative twitch and the sudden increase in current when the light stops, have also been observed by Minchin in photoelectric cells.
(1) The decline may nearly reach zero. If now the light be cut off there is a rebound towards recovery downwards, which carries it below zero, followed by an after-oscillation (fig. 108, a).
(1) The decline could almost hit zero. If the light is turned off now, there’s a bounce back towards recovery downwards, which takes it below zero, followed by a delayed oscillation (fig. 108, a).
(2) If the light be continued for a longer time, the decline goes on even below zero; that is to say, the response now becomes apparently negative. If, now, the light be stopped, there is a rebound upwards to recovery, with, generally speaking, a slight preliminary twitch downwards (fig. 108, b, c). This rebound carries it back, not only to the zero position, but sometimes beyond that position. We have here a parallel to the following observation of Dewar and McKendrick: ‘When diffuse light is allowed to impinge on the eye of the frog, after it has arrived at a tolerably stable condition, the natural E.M.F. is in the first place increased, then diminished; during the continuance of light it is still slowly diminished to a point where it remains tolerably constant, and on the removal of light there is a sudden increase of the E.M. power nearly up to its original position.’[18]
(2) If the light is kept on for a longer period, the decline continues even below zero; in other words, the response now seems to be negative. When the light is turned off, there's an upward rebound towards recovery, usually with a slight initial twitch downwards (fig. 108, b, c). This rebound brings it back not only to the zero position but sometimes past it. This is similar to the following observation by Dewar and McKendrick: ‘When diffuse light is allowed to hit the eye of the frog, after reaching a fairly stable condition, the natural E.M.F. first increases, then decreases; while the light continues, it slowly decreases to a point where it remains fairly constant, and upon removing the light, there's a sudden increase in the E.M. power almost back to its original position.’[18]

Fig. 108—Decline under the Continued Action of Light
Fig. 108—Decline Under Ongoing Exposure to Light
(a) Decline short of zero; on stoppage of light, rebound downwards to zero; after-oscillation.
(a) Drop just below zero; when the light stops, it falls back down to zero; then it overshoots a bit.
(b) Decline below zero; on stoppage of light, rebound towards zero, with preliminary negative twitch.
(b) Drop below zero; when the light stops, bounce back toward zero, with an initial negative twitch.
(c) The same, decline further down; negative twitch almost disappearing.
(c) The same, declining further down; the negative twitch is almost gone.
(3) I have sometimes obtained the following curious result. On the incidence of light there is a response, say, upward. On the continuation of light the response declines to zero and remains at the zero position, there being no further action during the continuation of stimulus. But on the cessation or ‘break’ of light stimulus, there is a response downwards, followed by the usual recovery. This reminds us of a somewhat similar responsive action produced by constant electric current on the muscle. At the moment of ‘make’ there is a responsive twitch, but afterwards the muscle remains quiescent during the passage of the current, but on breaking the current there is seen a second responsive twitch.
(3) I have sometimes observed the following interesting result. When light hits a surface, there's an upward reaction. As the light continues, this response gradually decreases to zero and stays there, with no further reaction while the light is on. However, when the light stops or breaks, there's a downward response, followed by the usual recovery. This is similar to the reaction produced by a constant electric current on a muscle. When the current is switched on, there’s an initial twitch, but then the muscle stays still while the current flows. When the current is turned off, another twitch is observed.
Résumé.—So we see that the response of the sensitive inorganic cell, to the stimulus of light, is in every way similar to that of the retina. In both we have, under normal conditions, a positive variation; in both the intensity of response up to a certain limit increases with the duration of illumination; it is affected, in both alike, by temperature; in both there is comparatively little fatigue; the increase of response with intensity of stimulus is similar in both; and finally, even in abnormalities—such as reversal of response, preliminary negative twitch on commencement, and terminal positive twitch on cessation of illumination, and decline and reversal under continued action of light—parallel effects are noticed.
Résumé.—So we can see that how the sensitive inorganic cell reacts to light is very similar to how the retina works. In both cases, under normal conditions, there's a positive change; in both, the strength of the response increases with the duration of light exposure, up to a certain limit; both are influenced by temperature; neither experiences much fatigue; the increase in response with the intensity of the stimulus is similar in both; and finally, even in unusual cases—like a change in response direction, an initial negative reaction when light starts, and a final positive reaction when light stops, along with decline and reversal when light is continuously applied—similar effects are observed.

We may notice here certain curious relations even in these abnormal responses (fig. 109). If the equilibrium position remain always constant, then it is easy to understand how, when the rising curve has attained its maximum, on the cessation of light, recovery should proceed downwards, towards the equilibrium position (fig. 109, a). One can also understand how, after reversal by the continued action of light, there should be a recovery upwards towards the old equilibrium position (fig. 109, b). What is curious is that in certain cases we get, on the stoppage of light, a preliminary twitch away from the zero or equilibrium position, upwards as in (c) (compare also fig. 107) and downwards as in (d) (compare also fig. 108 b).
We can see some interesting relationships even in these unusual responses (fig. 109). If the equilibrium position is always steady, then it's easy to understand why, when the rising curve reaches its peak and the light stops, the recovery should move downward towards the equilibrium position (fig. 109, a). It also makes sense that after being reversed by prolonged exposure to light, there would be a recovery upward towards the previous equilibrium position (fig. 109, b). What's interesting is that in some cases, when the light stops, we see an initial twitch away from the zero or equilibrium position, upward as in (c) (also see fig. 107) and downward as in (d) (also see fig. 108 b).
CHAPTER XIX
VISUAL ANALOGUES
- Effect of light of short duration
- —After-oscillation
- —Positive and negative after-images
- —Binocular alternation of vision
- —Period of alternation modified by physical condition
- —After-images and their revival
- —Unconscious visual impression.
We have already referred to the electrical theory of the visual impulse. We have seen how a flash of light causes a transitory electric impulse not only in the retina, but also in its inorganic substitute. Light thus produces not only a visual but also an electrical impulse, and it is not improbable that the two may be identical. Again, varying intensities of light give rise to corresponding intensities of current, and the curves which represent the relation between the increasing stimulus and the increasing response have a general agreement with the corresponding curve of visual sensation. In the present chapter we shall see how this electrical theory not only explains in a simple manner ordinary visual phenomena, but is also deeply suggestive with regard to others which are very obscure.
We’ve already talked about the electrical theory of visual perception. We’ve observed how a flash of light creates a temporary electric impulse not just in the retina, but also in its inorganic counterpart. Light, therefore, generates not just a visual but also an electrical impulse, and it’s quite possible that the two are the same. Moreover, different intensities of light lead to proportional intensities of current, and the graphs that show the relationship between the increasing stimulus and the increasing response align closely with the corresponding curve of visual sensation. In this chapter, we will explore how this electrical theory not only provides a straightforward explanation for ordinary visual phenomena but also offers insightful implications for other, more obscure experiences.
We have seen in our silver cell that if the molecular conditions of the anterior and posterior surfaces were exactly similar, there would be no current. In practice, however, this is seldom the case. There is, generally speaking, a slight difference, and a feeble current in the circuit. It is thus seen that there may be an existing feeble current, to which the effect of light is added algebraically. The stimulus of light may thus increase the existing current of darkness (positive variation). On the cessation of light again, the current of response disappears and there remains only the feeble original current.
We have observed in our silver cell that if the molecular conditions of the front and back surfaces were exactly the same, there would be no current. In reality, though, this is rarely the case. Generally speaking, there is a slight difference, leading to a weak current in the circuit. Thus, it’s clear that there can be a weak existing current, to which the effect of light adds up. The stimulus of light can increase the existing current created by darkness (positive change). When the light stops, the response current disappears, and only the weak original current remains.
In the case of the retina, also, it is curious to note that on closing the eye the sensation is not one of absolute darkness, but there is a general feeble sensation of light, known as ‘the intrinsic light of the retina.’ The effect produced by external light is superposed on this intrinsic light, and certain curious results of this algebraical summation will be noticed later.
In the case of the retina, it's also interesting to point out that when you close your eyes, the feeling isn't one of complete darkness, but rather a faint sensation of light, known as 'the intrinsic light of the retina.' The effect of external light is layered on top of this intrinsic light, and some intriguing outcomes of this combined effect will be discussed later.

Fig. 110—Response-curves of the Sensitive Silver Cell
Fig. 110—Response curves of the Sensitive Silver Cell
Showing greater persistence of after-effect when the stimulus is strong.
Showing more enduring after-effects when the stimulus is strong.
(a) Short exposure of 2″ to light of intensity 1; (b) short exposure of 2″ to light nine times as strong.
(a) Short exposure of 2″ to light with an intensity of 1; (b) short exposure of 2″ to light nine times stronger.
Effect of light of short duration.—If we subject the sensitive cell to a flash of radiation, the effect is not instantaneous but grows with time. It attains a maximum some little time after the incidence of light, and the effect then gradually passes away. Again, as we have seen previously with regard to mechanical strain, the after-effect persists for a slightly longer time when the stimulus is stronger. The same is true of the after-effect of the stimulus of light. Two curves which exhibit this are given below (fig. 110). With regard to the first point—that the maximum effect is attained some time after the cessation of a short exposure—the corresponding experiment on the eye may be made as follows: at the end of a tube is fixed a glass disc coated with lampblack, on which, by scratching with a pin, some words are written in transparent characters. The length of the tube is so adjusted that the disc is at the distance of most distinct vision from the end of the tube applied to the eye. The blackened disc is turned towards a source of strong light, and a short exposure is given by the release of a photographic shutter interposed between the disc and the eye. On closing the eye, immediately after a short exposure, it will at first be found that there is hardly any well-defined visual sensation; after a short time, however, the writing on the blackened disc begins to appear in luminous characters, attains a maximum intensity, and then fades away. In this case the stimulus is of short duration, the light being cut off before the maximum effect is attained. The after-effect here is positive, there being no reversal or interval of darkness between the direct image and the after-image, the one being merely the continuation of the other. But we shall see, if light is cut off after a maximum effect is attained by long exposure, that the immediate after-image would be negative (see below). The relative persistence of after-effect of lights of different intensities may be shown in the following manner:
Effect of short-duration light.—When we expose a sensitive cell to a flash of light, the effect doesn't happen immediately but gradually increases over time. It reaches a peak some time after the light hits and then slowly decreases. As we've mentioned before regarding mechanical strain, the after-effect lasts a bit longer when the stimulus is stronger. The same applies to the after-effect of light. Two curves that illustrate this are included below (fig. 110). To demonstrate the first point—that the maximum effect occurs some time after a brief exposure—we can conduct an experiment on the eye as follows: At the end of a tube, there is a glass disc covered in black paint, on which some words are scratched in transparent characters with a pin. The tube is adjusted so that the disc is at the distance where the vision is clearest. The blackened disc faces a strong light source, and a brief exposure is provided by activating a photographic shutter placed between the disc and the eye. After closing the eye immediately after this short exposure, there will initially be almost no clear visual sensation; however, after a moment, the writing on the blackened disc starts to appear in bright characters, reaching maximum intensity before fading away. In this scenario, the stimulus lasts a short time, with the light being turned off before the maximum effect is reached. The after-effect in this case is positive, without any reversal or interval of darkness between the direct image and the after-image; one simply continues from the other. However, we will see that if the light is turned off after reaching maximum effect due to a longer exposure, the immediate after-image will be negative (see below). The relative persistence of after-effects from lights of varying intensities can be demonstrated as follows:
If a bold design be traced with magnesium powder on a blackened board and fired in a dark room, the observer not being acquainted with the design, the instantaneous flash of light, besides being too quick for detailed observation, is obscured by the accompanying smoke. But if the eyes be closed immediately after the flash, the feebler obscuring sensation of smoke will first disappear, and will leave clear the more persistent after-sensation of the design, which can then be read distinctly. In this manner I have often been able to see distinctly, on closing the eyes, extremely brief phenomena of light which could not otherwise have been observed, owing either to their excessive rapidity or to their dazzling character.[19]
If you trace a bold design with magnesium powder on a black board and light it in a dark room, anyone who isn't familiar with the design will see an instant flash of light that is too quick to study in detail and is also hidden by the smoke that follows. However, if you close your eyes right after the flash, the lighter smoke will fade away first, leaving the more lasting after-image of the design which can then be seen clearly. This way, I have often been able to observe very brief light phenomena when I close my eyes, which wouldn’t be noticed otherwise due to their quickness or brightness.[19]
After-oscillation.—In the case of the sensitive silver cell, we have seen (fig. 105), when it has been subjected for some time to strong light, that the current of response attains a maximum, and that on the stoppage of the stimulus there is an immediate rebound towards recovery. In this rebound there may be an over-shooting of the equilibrium position, and an after-oscillation is thus produced.
After-oscillation.—With the sensitive silver cell, we have observed (fig. 105) that after being exposed to strong light for a while, the response current reaches its peak. When the light is turned off, there's a quick rebound as it tries to return to normal. This rebound can sometimes overshoot the balance point, resulting in an after-oscillation.
Visual recurrence.—Translated into the visual circuit, this would mean an alternating series of after-images. On the cessation of light of strong intensity and long duration, the immediate effect would be a negative rebound, unlike the positive after-effect which followed on a short exposure.
Visual recurrence.—In visual terms, this would involve a series of alternating afterimages. When strong light is turned off after a long exposure, the immediate response would be a negative rebound, unlike the positive after-effect that happens after a brief exposure.
The next rebound is positive, giving rise to a sensation of brightness. This will go on in a recurrent series.
The next bounce back is positive, creating a feeling of brightness. This will continue in a repeated pattern.
If we look for some time at a very bright object, preferably with one eye, on closing the eye there is an immediate dark sensation followed by a sensation of light. These go on alternating and give rise to the phenomena of recurrent vision. With the eyes closed, the positive or luminous phases are the more prominent.
If we stare at a really bright object for a while, especially with one eye, when we close that eye, we first feel a quick darkness followed by a sense of light. These sensations keep alternating and create the experience of recurring vision. With our eyes closed, the bright or light phases stand out more.
This phenomenon may be observed in a somewhat different manner. After staring at a bright light we may look towards a well-lighted wall. The dark phases will now become the more noticeable.
This phenomenon can be seen in a slightly different way. After staring at a bright light, we might look at a brightly lit wall. The dark areas will now stand out more.
If, however, we look towards a dimly lighted wall, both the dark and bright phases will be noticed alternately.
If we look at a dimly lit wall, we will see the dark and bright areas alternate.
The negative effect is usually explained as due to fatigue. That position of the retina affected by light is supposed to be ‘tired,’ and a negative image to be formed in consequence of exhaustion. By this exhaus tion is meant either the presence of fatigue-stuffs, or the breaking-down of the sensitive element of the tissue, or both of these. In such a case we should expect that this fatigue, with its consequent negative image, would gradually and finally disappear on the restoration of the retina to its normal condition.
The negative effect is usually explained as being caused by fatigue. That area of the retina affected by light is thought to be 'tired,' leading to a negative image forming due to exhaustion. By this exhaustion, we mean either the buildup of fatigue substances or the deterioration of the sensitive tissue element, or both. In this situation, we would expect that this fatigue, along with its resulting negative image, would gradually and ultimately fade away once the retina returns to its normal state.
We find, however, that this is not the case, for the negative image recurs with alternate positive. The accepted theory of fatigue is incapable of explaining this phenomenon.
We find, however, that this isn't true, as the negative image reappears alongside a positive one. The accepted theory of fatigue can't explain this phenomenon.
In the sensitive silver cell, we found that the molecular strain produced by light gave rise to a current of response, and that on the cessation of light an oscillatory after-effect was produced. The alternating after-effect in the retina points to an exactly similar process.
In the delicate silver cell, we discovered that the molecular stress created by light generated a responsive current, and when the light stopped, an oscillating after-effect occurred. The fluctuating after-effect in the retina indicates a very similar process.
Binocular alternation of vision.—It was while experimenting on the phenomena of recurrent vision that I discovered the curious fact that in normal eyes the two do not see equally well at a given instant, but that the visual effect in each eye undergoes fluctuation from moment to moment, in such a way that the sensation in the one is complementary to that in the other, the sum of the two sensations remaining approximately constant. Thus they take up the work of seeing, and then, relatively speaking, resting, alternately. This division of labour, in binocular vision, is of obvious advantage.
Binocular alternation of vision.—While experimenting with the phenomena of recurring vision, I discovered an interesting fact: in normal eyes, the two do not see equally well at the same time. Instead, the visual effect in each eye fluctuates from moment to moment, so that the sensation in one eye complements the sensation in the other, keeping the total of the two sensations roughly constant. Thus, they alternate between doing the work of seeing and, relatively speaking, resting. This division of labor in binocular vision is clearly beneficial.

As regards maximum sensation in the two retinæ there is then a relative retardation of half a period. This may be seen by means of a stereoscope, carrying, instead of stereo-photographs, incised plates through which we look at light. The design consists of two slanting cuts at a suitable distance from each other. One cut, R, slants to the right, and the other, L, to the left (see fig. 111). When the design is looked at through the stereoscope, the right eye will see, say R, and the left L, the two images will appear superimposed, and we see an inclined cross. When the stereoscope is turned towards the sky, and the cross looked at steadily for some time, it will be found, owing to the alternation already referred to, that while one arm of the cross begins to be dim, the other becomes bright, and vice versa. The alternate fluctuations become far more conspicuous when the eyes are closed; the pure oscillatory after-effects are then obtained in a most vivid manner. After looking through the stereoscope for ten seconds or more, the eyes are closed. The first effect observed is one of darkness, due to the rebound. Then one luminous arm of the cross first projects aslant the dark field, and then slowly disappears, after which the second (perceived by the other eye) shoots out suddenly in a direction athwart the first. This alternation proceeds for a long time, and produces the curious effect of two luminous blades crossing and recrossing each other.
When it comes to maximum sensation in both retinas, there’s a delay of half a period. You can see this using a stereoscope, which uses cut plates instead of stereo photographs to look at light. The design has two angled cuts spaced appropriately apart. One cut, R, angles right, and the other, L, angles left (see fig. 111). When you look at this design through the stereoscope, your right eye sees R and your left eye sees L, causing the two images to overlap and creating an inclined cross. If you tilt the stereoscope toward the sky and focus on the cross for a while, you’ll notice that because of the previously mentioned alternation, one arm of the cross starts to fade while the other brightens, and vice versa. These alternating fluctuations become more noticeable when your eyes are closed; the pure oscillatory aftereffects become very vivid. After looking through the stereoscope for around ten seconds or more, close your eyes. The first thing you’ll notice is darkness from the rebound. Then one shining arm of the cross will emerge diagonally across the dark field and then gradually fade, after which the second arm (seen by the other eye) will suddenly appear crossing the first. This back-and-forth continues for a long time, creating the unusual effect of two glowing blades crossing and re-crossing each other.
Another method of bringing out the phenomenon of alternation in a still more striking manner is to look at two different sets of writing, with the two eyes. The resultant effect is a blur, due to superposition, and the inscription cannot be read with the eyes open. But on closing them, the composite image is analysed alternately into its component parts, and thus we are enabled to read better with eyes shut than open.
Another way to highlight the phenomenon of alternation in an even more noticeable way is to look at two different sets of writing with each eye. The result is a blur, because of superposition, and the text can't be read with the eyes open. However, by closing them, the combined image is alternately analyzed into its separate parts, allowing us to read better with our eyes closed than open.
This period of alternation is modified by age and by the condition of the eye. It is, generally speaking, shorter in youth. I have seen it vary in different individuals from 1″ to 10″ or more. About 4″ is the most usual. With the same individual, again, the period is somewhat modified by previous conditions of rest or activity. Very early in the morning, after sleep, it is at its shortest. I give below a set of readings given by an observer:
This period of alternation changes with age and the state of the eye. Generally speaking, it's shorter in younger people. I've seen it vary among different individuals from 1" to 10" or more. About 4" is the most common. For the same person, the period can also be slightly influenced by prior states of rest or activity. Early in the morning, right after sleep, it’s at its shortest. Below, I provide a set of readings taken by an observer:
| Period | Period | ||||
|---|---|---|---|---|---|
| 8 | a.m. | 3″ | 6 | p.m. | 5·4″ |
| 12 | noon | 4″ | 9 | " | 5·6″ |
| 3 | p.m. | 5″ | 11 | " | 6·5″ |
Again, if one eye be cooled and the other warmed, the retinal oscillation in one eye is quicker than in the other. The quicker oscillation overtakes the slower, and we obtain the curious phenomenon of ‘visual beats.’
Again, if one eye is cooled and the other is warmed, the retinal oscillation in one eye is faster than in the other. The faster oscillation catches up to the slower one, and we get the interesting phenomenon of ‘visual beats.’
After-images and their revival.—In the experiment with the stereoscope and the design of the cross, the after-images of the cross seen with the eyes closed are at first very distinct—so distinct that any unevenness at the edges of the slanting cuts in the design can be distinctly made out. There can thus be no doubt of the ‘objective’ nature of the strain impression on the retina, which on the cessation of direct stimulus of light gives rise to after-oscillation with the concomitant visual recurrence. This recurrence may therefore be taken as a proof of the physical strain produced on the retina. The recurrent after-image is very distinct at the beginning and becomes fainter at each repetition; a time comes when it is difficult to tell whether the image seen is the objective after-effect due to strain or merely an effect of ‘memory.’ In fact there is no line of demarcation between the two, one simply merges into the other. That this ‘memory’ image is due to objective strain is rendered evident by its recurrence.
After-images and their revival.—In the experiment with the stereoscope and the design of the cross, the after-images of the cross seen with eyes closed are initially very clear—so clear that any unevenness at the edges of the slanted cuts in the design can be easily identified. This leaves no doubt about the ‘objective’ nature of the strain impression on the retina, which, upon the stopping of direct light stimulus, leads to after-oscillation along with the accompanying visual recurrence. Therefore, this recurrence can be considered proof of the physical strain exerted on the retina. The recurrent after-image is very vivid at first and becomes fainter with each repetition; eventually, it becomes hard to determine whether the image seen is the objective after-effect due to strain or simply a ‘memory’ effect. In fact, there is no clear line between the two; one simply blends into the other. The fact that this ‘memory’ image arises from objective strain is made clear by its recurrence.
In connection with this it is interesting to note that some of the undoubted phenomena of memory are also recurrent. ‘Certain sensations for which there is no corresponding process outside the body are generally grouped for convenience under this term [memory]. If the eyes be closed and a picture be called to memory, it will be found that the picture cannot be held, but will repeatedly disappear and appear.’[20]
In relation to this, it's interesting to point out that some clear memory phenomena are also recurring. "Certain sensations that don’t have a corresponding process outside the body are usually categorized under this term [memory]. If your eyes are closed and you try to recall a picture, you’ll find that you can’t hold onto the image; it will repeatedly fade in and out."[20]
The visual impressions and their recurrence often persist for a very long time. It usually happens that owing to weariness the recurrent images disappear; but in some instances, long after this disappearance, they will spontaneously reappear at most unexpected moments. In one instance the recurrence was observed in a dream, about three weeks after the original impression was made. In connection with this, the revival of images, on closing the eyes at night, that have been seen during the day, is extremely interesting.
The visual impressions and their recurrence often last for a long time. Usually, due to fatigue, the recurring images fade away; however, in some cases, long after they disappear, they can unexpectedly return. In one case, the recurrence was noted in a dream about three weeks after the initial impression was made. Related to this, it’s really interesting how images seen during the day can revive when you close your eyes at night.
Unconscious visual impression.—While repeating certain experiments on recurrent vision, the above phenomenon became prominent in an unexpected manner. I had been intently looking at a particular window, and obtaining the subsequent after-images by closing the eye; my attention was concentrated on the window, and I saw nothing but the window either as a direct or as an after effect. After this had been repeated a number of times, I found on one occasion, after closing the eye, that, owing to weariness of the particular portion of the retina, I could no longer see the after-image of the window; instead of this I however saw distinctly a circular opening closed with glass panes, and I noticed even the jagged edges of a broken pane. I was not aware of the existence of a circular opening higher up in the wall. The image of this had impressed itself on the retina without my knowledge, and had undoubtedly been producing the recurrent images which remained unnoticed because my principal field of after-vision was filled up and my attention directed towards the recurrent image of the window. When this failed to appear, my field of after-vision was relatively free from distraction, and I could not help seeing what was unnoticed before. It thus appears that, in addition to the images impressed in the retina of which we are conscious, there are many others which are imprinted without our knowledge. We fail to notice them because our attention is directed to something else. But at a subsequent period, when the mind is in a passive state, these impressions may suddenly revive owing to the phenomenon of recurrence. This observation may afford an explanation of some of the phenomena connected with ocular phantoms and hallucinations not traceable to any disease. In these cases the psychical effects produced appear to have no objective cause. Bearing in mind the numerous visual impressions which are being unconsciously made on the retina, it is not at all unlikely that many of these visual phantoms may be due to objective causes.
Unconscious visual impression.—While repeating certain experiments on recurring vision, this phenomenon became obvious in an unexpected manner. I had been intensely staring at a specific window and getting after-images by closing my eye; my focus was solely on the window, and I saw nothing else, either directly or as an aftereffect. After doing this several times, I discovered on one occasion that, due to fatigue in that part of my retina, I could no longer see the after-image of the window. Instead, I distinctly saw a circular opening with glass panes, and I even noticed the jagged edges of a broken pane. I wasn't aware of the existence of a circular opening higher up on the wall. The image of this had imprinted itself on my retina without me knowing, and it was likely responsible for the recurring images that went unnoticed because my main focus was on the recurring image of the window. When that image didn't appear, my after-vision field was relatively clear of distractions, allowing me to see what I had previously overlooked. It seems that, in addition to the images we consciously recognize on the retina, there are many others imprinted without our awareness. We don't notice them because our attention is on something else. However, later on, when the mind is more passive, these impressions can suddenly resurface due to recurrence. This observation might explain some phenomena related to visual phantoms and hallucinations that can't be linked to any illness. In these cases, the psychological effects seem to lack an objective cause. Considering the numerous visual impressions being made unconsciously on the retina, it's quite possible that many of these visual phantoms may stem from objective causes.
FOOTNOTES:
[19] As an instance of this I may mention the experiment which I saw on the quick fusion of metals exhibited at the Royal Institution by Sir William Roberts-Austen (1901), where, owing to the glare and the dense fumes, it was impossible to see what happened in the crucible. But I was able to see every detail on closing the eyes. The effects of the smoke, being of less luminescence, cleared away first, and left the after-image of the molten metal growing clearer on the retina.
[19] One example I can mention is the experiment I witnessed on the rapid fusion of metals at the Royal Institution by Sir William Roberts-Austen (1901). Because of the bright light and heavy fumes, it was impossible to see what was happening in the crucible. However, I was able to see every detail when I closed my eyes. The smoke, being less bright, cleared away first, revealing a clearer after-image of the molten metal on my retina.
CHAPTER XX
GENERAL SURVEY AND CONCLUSION
We have seen that stimulus produces a certain excitatory change in living substances, and that the excitation produced sometimes expresses itself in a visible change of form, as seen in muscle; that in many other cases, however—as in nerve or retina—there is no visible alteration, but the disturbance produced by the stimulus exhibits itself in certain electrical changes, and that whereas the mechanical mode of response is limited in its application, this electrical form is universal.
We have observed that a stimulus causes a specific type of excitement in living things, and that this excitement can sometimes show up as a visible change in shape, like in muscles. However, in many other cases—such as with nerves or the retina—there isn't any visible change, but the disruption caused by the stimulus is reflected in certain electrical changes. While the mechanical way of responding is limited in its use, this electrical response is universal.
This irritability of the tissue, as shown in its capacity for response, electrical or mechanical, was found to depend on its physiological activity. Under certain conditions it could be converted from the responsive to an irresponsive state, either temporarily as by anæsthetics, or permanently as by poisons. When thus made permanently irresponsive by any means, the tissue was said to have been killed. We have seen further that from this observed fact—that a tissue when killed passes out of the state of responsiveness into that of irresponsiveness; and from a confusion of ‘dead’ things with inanimate matter, it has been tacitly assumed that inorganic substances, like dead animal tissues, must necessarily be irresponsive, or incapable of being excited by stimulus—an assumption which has been shown to be gratuitous.
This irritation of the tissue, as evidenced by its ability to respond, whether electrically or mechanically, was found to depend on its physiological activity. Under certain conditions, it could switch from a responsive to an unresponsive state, either temporarily through anesthetics or permanently through poisons. When it was permanently made unresponsive by any means, the tissue was said to have been killed. We have further observed that from this fact—that a tissue becomes unresponsive when killed—and from confusing ‘dead’ things with inanimate matter, it has been implicitly assumed that inorganic substances, like dead animal tissues, must also be unresponsive or incapable of being stimulated—an assumption that has proven to be unfounded.
This ‘unexplained conception of irritability became the starting-point,’ to quote the words of Verworn,[21] ‘of vitalism, which in its most complete form asserted a dualism of living and lifeless Nature.... The vitalists soon,’ as he goes on to say, ‘laid aside, more or less completely, mechanical and chemical explanations of vital phenomena, and introduced, as an explanatory principle, an all-controlling unknown and inscrutable “force hypermécanique.” While chemical and physical forces are responsible for all phenomena in lifeless bodies, in living organisms this special force induces and rules all vital actions.
This "unexplained idea of irritability became the starting point," to quote Verworn, [21] "of vitalism, which in its most complete form claimed a dualism between living and non-living nature.... The vitalists soon," as he continues, "mostly set aside mechanical and chemical explanations for vital phenomena and introduced an all-controlling unknown and mysterious 'hypermécanique force' as an explanatory principle. While chemical and physical forces explain all phenomena in non-living bodies, in living organisms this special force governs and drives all vital actions."
‘Later vitalists, however, attempted no analysis of vital force; they employed it in a wholly mystical form as a convenient explanation of all sorts of vital phenomena.... In place of a real explanation a simple phrase such as “vital force” was satisfactory, and signified a mystical force belonging to organisms only. Thus it was easy to “explain” the most complex vital phenomena.’
‘Later vitalists, however, didn’t analyze vital force; they used it in a completely mystical way as an easy explanation for all kinds of vital phenomena.... Instead of a real explanation, a simple phrase like “vital force” was enough and signified a mystical power unique to living organisms. This made it simple to “explain” even the most complex vital phenomena.’
From this position, with its assumption of the super-physical character of response, it is clear that on the discovery of similar effects amongst inorganic substances, the necessity of theoretically maintaining such dualism in Nature must immediately fall to the ground.
From this stance, with its assumption of the non-physical nature of response, it's evident that upon discovering similar effects in inorganic substances, the need to theoretically uphold such dualism in Nature must instantly collapse.
In the previous chapters I have shown that not the fact of response alone, but all those modifications in response which occur under various conditions, take place in plants and metals just as in animal tissues. It may now be well to make a general survey of these phenomena, as exhibited in the three classes of substances.
In the earlier chapters, I demonstrated that it's not just the act of responding that matters, but also all the changes in response that happen under different conditions, which occur in plants and metals just like they do in animal tissues. It’s a good idea to take a general look at these phenomena as they appear in the three types of substances.
We have seen that the wave of molecular disturbance in a living animal tissue under stimulus is accompanied by a wave of electrical disturbance; that in certain types of tissue the stimulated is relatively positive to the less disturbed, while in others it is the reverse; that it is essential to the obtaining of electric response to have the contacts leading to the galvanometer unequally affected by excitation; and finally that this is accomplished either (1) by ‘injuring’ one contact, so that the excitation produced there would be relatively feeble, or (2) by introducing a perfect block between the two contacts, so that the excitation reaches one and not the other.
We’ve observed that when an animal tissue is stimulated, it experiences a wave of molecular activity along with a wave of electrical activity. In some types of tissue, the stimulated area is relatively more positive than the less stimulated area, while in other tissues, it’s the opposite. To get an electrical response, it’s crucial that the contacts leading to the galvanometer are affected differently by the excitation. This can be achieved either (1) by ‘damaging’ one contact so that the excitation there is weaker, or (2) by creating a complete block between the two contacts, allowing the excitation to reach one and not the other.
Further, it has been shown that this characteristic of exhibiting electrical response under stimulus is not confined to animal, but extends also to vegetable tissues. In these the same electrical variations as in nerve and muscle were obtained, by using the method of injury, or that of the block.
Further, it has been shown that this characteristic of showing electrical response under stimulus is not just limited to animals, but also extends to plant tissues. In these cases, the same electrical variations seen in nerves and muscles were obtained using the method of injury or the block method.
Passing to inorganic substances, and using similar experimental arrangements, we have found the same electrical responses evoked in metals under stimulus.
Passing to inorganic substances, and using similar experimental setups, we have found the same electrical responses triggered in metals under stimulation.
Negative variation.—In all cases, animal, vegetable, and metal, we may obtain response by the method of negative variation, so called, by reducing the excitability of one contact by physical or chemical means. Stimulus causes a transient diminution of the existing current, the variation depending on the intensity of the stimulus (figs. 4, 7, 54).
Negative variation.—In all cases involving animals, plants, and metals, we can achieve a response through the method of negative variation, which involves decreasing the excitability of one contact using physical or chemical methods. The stimulus results in a temporary reduction of the existing current, with the variation depending on the strength of the stimulus (figs. 4, 7, 54).

Fig. 112.—Uniform Responses in (A) Nerve, (P) Plant, and (M) Metal
Fig. 112.—Consistent Reactions in (A) Nerve, (P) Plant, and (M) Metal
The normal response in nerve is represented ‘down.’ In this and following figures, (A) is the record of responses in animal, (P) in plant, and (M) in metal.
The normal response in nerves is shown as ‘down.’ In this and the following figures, (A) shows the record of responses in animals, (P) in plants, and (M) in metal.
Relation between stimulus and response.—In all three classes we have found that the intensity of response increases with increasing stimulus. At very high intensities of stimulus, however, there is a tendency of the response to reach a limit (figs. 30, 32, 84). The law that is known as Weber-Fechner’s shows a similar characteristic in the relation between stimulus and sensation. And if sensation be a measure of physiological effect we can understand this correspondence of the physiological and sensation curves. We now see further that the physiological effects themselves are ultimately reducible to simple physical phenomena.
Relation between stimulus and response.—In all three classes, we have noticed that the intensity of the response increases as the stimulus gets stronger. However, at very high levels of stimulus intensity, the response tends to plateau (figs. 30, 32, 84). The principle known as Weber-Fechner’s illustrates a similar pattern in the relationship between stimulus and sensation. If we consider sensation as a measure of physiological effect, it helps us understand the relationship between the physiological and sensation curves. We now also see that the physiological effects can ultimately be broken down into basic physical phenomena.
Effects of superposition.—In all three types, ineffective stimuli become effective by superposition.
Effects of superposition.—In all three types, ineffective stimuli become effective through superposition.
Uniform responses.—In the responses of animal, vegetable, and metal alike we meet with a type where the responses are uniform (fig. 112).
Uniform responses.—In the responses of animals, plants, and metals, we encounter a type where the responses are consistent (fig. 112).

Fig. 113.—Fatigue (A) in Muscle, (P) in Plant, (M) in Metal
Fig. 113.—Fatigue (A) in Muscle, (P) in Plant, (M) in Metal
The explanation hitherto given of fatigue in animal tissues—that it is due to dissimilation or breakdown of tissue, complicated by the presence of fatigue-products, while recovery is due to assimilation, for which material is brought by the blood-supply—has long been seen to be inadequate, since the restorative effect succeeds a short period of rest even in excised bloodless muscle. But that the phenomena of fatigue and recovery were not primarily dependent on dissimilation or assimilation becomes self-evident when we find exactly similar effects produced not only in plants, but also in metals (fig. 113). It has been shown, on the other hand, that these effects are primarily due to cumulative residual strains, and that a brief period of rest, by removing the overstrain, removes also the sign of fatigue.
The explanation so far provided about fatigue in animal tissues—that it results from the breakdown of tissue, complicated by the presence of fatigue products, while recovery comes from the process of assimilation, with materials supplied by the blood—has long been considered inadequate. This is because the recovery effect happens after just a short rest period, even in excised bloodless muscle. However, it becomes obvious that the experiences of fatigue and recovery are not primarily based on breakdown or assimilation when we see similar effects occurring not just in plants but also in metals (fig. 113). On the other hand, it has been demonstrated that these effects mainly come from cumulative residual strains, and that a brief rest period helps remove the overstrain, thus eliminating the signs of fatigue.
Staircase effect.—The theory of dissimilation due to stimulus reducing the functional activity below par, and thus causing fatigue, is directly negatived by what is known as the ‘staircase’ effect, where successive equal stimuli produce increasing response. We saw an exactly similar phenomenon in plants and metals, where successive responses to equal stimuli exhibited an increase, apparently by a gradual removal of molecular sluggishness (fig. 114).
Staircase effect.—The theory of dissimilation, which suggests that stimulus reduces functional activity below normal levels and causes fatigue, is directly contradicted by what’s called the ‘staircase’ effect, where repeated equal stimuli lead to an increased response. We observed an exactly similar phenomenon in plants and metals, where repeated responses to equal stimuli showed an increase, seemingly due to a gradual removal of molecular sluggishness (fig. 114).

Increased response after continuous stimulation.—An effect somewhat similar, that is to say, an increased response, due to increased molecular mobility, is also shown sometimes after continuous stimulation, not only in animal tissues, but also in metals (fig. 115).
Increased response after continuous stimulation.—A similar effect, meaning an increased response, caused by increased molecular mobility, is sometimes observed after continuous stimulation, not just in animal tissues, but also in metals (fig. 115).
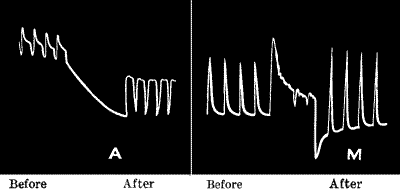
Fig. 115.—Increased Response after Continuous Stimulation in Nerve and Metal
Fig. 115.—Enhanced Response after Ongoing Stimulation in Nerve and Metal
The normal response in animal tissue is represented ‘down,’ in metal ‘up.’
The typical response in animal tissue is shown as 'down,' while in metal it's 'up.'
Modified response.—In the case of nerve we saw that the normal response, which is negative, sometimes becomes reversed in sign, i.e. positive, when the specimen is stale. In retina again the normal positive response is converted into negative under the same conditions. Similarly, we found that a plant when withering often shows a positive instead of the usual negative response (fig. 28). On nearing the death-point, also by subjection to extremes of temperature, the same reversal of response is occasionally observed in plants. This reversal of response due to peculiar molecular modification was also seen in metals.
Modified response.—In the case of nerve response, we observed that the typical reaction, which is negative, can sometimes switch to positive when the specimen is old. Similarly, in the retina, the usual positive response turns into a negative one under the same conditions. We also noted that a wilting plant often exhibits a positive response instead of the typical negative one (fig. 28). Approaching the death-point and when exposed to extreme temperatures, we occasionally see this same reversal of response in plants. This reversal of response due to unusual molecular changes was also observed in metals.

Fig. 116.—Modified Abnormal Response in (A) Nerve and (M) Metal converted into Normal, after Continuous Stimulation
Fig. 116.—Altered Abnormal Response in (A) Nerve and (M) Metal changed to Normal, after Ongoing Stimulation
(A) is the record for nerve (recording galvanometer not being dead-beat shows after-oscillation); the abnormal ‘up’ is converted into normal ‘down’ after continuous stimulation. (M) is the record for metal, the abnormal ‘down’ being converted into normal ‘up’ after like stimulation.
(A) is the record for nerve (the recording galvanometer, which isn't dead-beat, shows after-oscillation); the unusual 'up' signal is turned into a normal 'down' after continuous stimulation. (M) is the record for metal, where the unusual 'down' signal is turned into a normal 'up' after similar stimulation.
But these modified responses usually become normal when the specimen is subjected to stimulation either strong or long continued (fig. 116).
But these modified responses typically become normal when the specimen is exposed to either strong or prolonged stimulation (fig. 116).
Diphasic variation.—A diphasic variation is observed in nerve, if the wave of molecular disturbance does not reach the two contacts at the same moment, or if the rate of excitation is not the same at the two points. A similar diphasic variation is also observed in the responses of plants and metals (figs. 26, 68).
Effect of temperature.—In animal tissues response becomes feeble at low temperatures. At an optimum temperature it reaches its greatest amplitude, and, again, beyond a maximum temperature it is very much reduced.
Effect of temperature.—In animal tissues, the response becomes weak at low temperatures. It reaches its peak at an ideal temperature, and then decreases significantly beyond a maximum temperature.
Effect of chemical reagents.—Finally, just as the response of animal tissue is exalted by stimulants, lowered by depressants, and abolished by poisons, so also we have found the response in plants and metals undergoing similar exaltation, depression, or abolition.
Effect of chemical reagents.—Finally, just as the reaction of animal tissue is enhanced by stimulants, reduced by depressants, and eliminated by poisons, we have also discovered that the response in plants and metals undergoes similar enhancement, reduction, or elimination.
We have seen that the criterion by which vital response is differentiated is its abolition by the action of certain reagents—the so-called poisons. We find, however, that ‘poisons’ also abolish the responses in plants and metals (fig. 117). Just as animal tissues pass from a state of responsiveness while living to a state of irresponsiveness when killed by poisons, so also we find metals transformed from a responsive to an irresponsive condition by the action of similar ‘poisonous’ reagents.
We’ve noticed that what distinguishes vital response is that it’s eliminated by the action of specific substances—what we refer to as poisons. However, we observe that these ‘poisons’ also eliminate responses in plants and metals (fig. 117). Just as animal tissues change from being responsive when alive to becoming unresponsive when killed by poisons, metals also transition from a responsive to an unresponsive state through the action of similar ‘poisonous’ substances.
The parallel is the more striking since it has long been known with regard to animal tissues that the same drug, administered in large or small doses, might have opposite effects, and in preceding chapters we have seen that the same statement holds good of plants and metals also.
The comparison is even more noticeable because it has been well established that with animal tissues, the same drug given in either large or small doses can produce opposite effects. In earlier chapters, we have found that this is also true for plants and metals.
Stimulus of light.—Even the responses of such a highly specialised organ as the retina are strictly paralleled by inorganic responses. We have seen how the stimulus of light evokes in the artificial retina responses which coincide in all their detail with those produced in the real retina. This was seen in ineffective stimuli becoming effective after repetition, in the relation between stimulus and response, and in the effects produced by temperature; also in the phenomenon of after-oscillation. These similarities went even further, the very abnormalities of retinal response finding their reflection in the inorganic.
Light stimulation.—Even the reactions of a highly specialized organ like the retina are closely matched by inorganic reactions. We've observed how light stimulates the artificial retina to produce responses that mirror exactly those of the real retina. This was evident in how ineffective stimuli became effective after being repeated, in the relationship between stimulus and response, and in the effects caused by temperature; it was also seen in the phenomenon of after-oscillation. These similarities extended even further, with the unusual reactions of the retina being reflected in the inorganic.

Fig. 117.—Abolition of Response in Nerve, Plant, and Metal by the Action of the same ‘Poison’
Fig. 117.—End of Response in Nerve, Plant, and Metal Due to the Effect of the Same 'Poison'
The first half in each set shows the normal response, the second half the abolition of response after the application of the reagent.
The first half of each set shows the normal response, while the second half shows the lack of response after applying the reagent.
Thus living response in all its diverse manifestations is found to be only a repetition of responses seen in the inorganic. There is in it no element of mystery or caprice, such as we must admit to be applied in the assumption of a hypermechanical vital force, acting in contradiction or defiance of those physical laws that govern the world of matter. Nowhere in the entire range of these response-phenomena—inclusive as that is of metals, plants, and animals—do we detect any breach of continuity. In the study of processes apparently so complex as those of irritability, we must, of course, expect to be confronted with many difficulties. But if these are to be overcome, they, like others, must be faced, and their investigation patiently pursued, without the postulation of special forces whose convenient property it is to meet all emergencies in virtue of their vagueness. If, at least, we are ever to understand the intricate mechanism of the animal machine, it will be granted that we must cease to evade the problems it presents by the use of mere phrases which really explain nothing.
Living responses, in all their different forms, are really just repetitions of responses seen in non-living things. There's no mystery or randomness in it, unlike what we might consider with the idea of a hypermechanical vital force that operates against the physical laws that control the material world. Throughout all these response phenomena—including metals, plants, and animals—we find no breaks in continuity. Studying processes that seem as complex as irritability will clearly present us with challenges. But if we want to overcome them, we need to confront these challenges directly and investigate them thoroughly, without assuming special forces that conveniently address all issues due to their ambiguity. If we ever want to truly understand the complex workings of the animal machine, we must stop dodging the issues it raises by using terms that ultimately explain nothing.
We have seen that amongst the phenomena of response, there is no necessity for the assumption of vital force. They are, on the contrary, physico-chemical phenomena, susceptible of a physical inquiry as definite as any other in inorganic regions.
We have seen that among the phenomena of response, there is no need to assume the existence of a vital force. They are, instead, physico-chemical phenomena that can be investigated with the same precision as any other phenomena in inorganic fields.
Physiologists have taught us to read in the response-curves a history of the influence of various external agencies and conditions on the phenomenon of life. By these means we are able to trace the gradual diminution of responsiveness by fatigue, by extremes of heat and cold, its exaltation by stimulants, the arrest of the life-process by poison.
Physiologists have shown us how to interpret response curves to understand how different external factors and conditions affect living organisms. This allows us to see how responsiveness gradually decreases due to fatigue and extreme temperatures, increases with stimulants, and is halted by poison.
The investigations which have just been described may possibly carry us one step further, proving to us that these things are determined, not by the play of an unknowable and arbitrary vital force, but by the working of laws that know no change, acting equally and uniformly throughout the organic and the inorganic worlds.
INDEX
| A | B | C | D | E | F | G | H | I | J | K | L | M |
| N | O | P | Q | R | S | T | U | V | W | X | Y | Z |
- Action current
- After-images and their revival, 177
- After-oscillation in photo-sensitive cell, 159, 163
- Anæsthetics, effect on response
- Annealing, effect on response in metal, 101, 138
- Chloral, effect on plant response, 75
- Chloroform, effect on
- Compensator, 22
- Current of injury in nerve, 7
- Curves, characteristics of response, 3
- Death-point, determination of, in plants, 61, 63
- Depressants, effect on inorganic response, 142
- Depression, response by relative, 87
- Dewar on retinal current, 149
- Diphasic variation
- Dose, effect on
- Electrical recorder, 11
- Electrical response. See Response, electrical
- Electric tapper, 24
- Exaltation, response by relative, 89
- Fatigue,
- absence of, under certain conditions,
- apparent, with increased frequency of stimulation,
- diminution of response under strong stimulus due to, in plant, 57
- in metal, 118, 119, 185
- in muscle, 118, 185
- in plant, 20, 185
- due to overstrain, 41
- rapid, under continuous stimulation
- removal of, by rest in plant, 43
- theory of, in muscle, 38, 185
- Injury, current of, in nerve, 7
- Inorganic response. See Metal, electrical response in
- Light,
- after-effect of short exposure to,
- decline and reversal of response under continuous,
- effect of temperature on
- relation between intensity and response to,
- response to,
- McKendrick on retinal response, 149
- Mechanical
- Metal, electric response in,
- abnormal, 125
- abolition of, by ‘poison,’ 143
- additive effect of superposition of stimulus on, 135
- annealing, effect of, on, 101
- by method of
- depressants, effect of, on, 142
- diphasic, 113, 114, 115, 116, 188
- enhancement of, after continuous stimulation, 127, 128, 186
- fatigue, 118, 119, 120, 121, 185. See also Fatigue
- maximum effect due to superposition of stimuli, 136
- modified, 129
- ‘molecular arrest,’ effect of, by ‘poison’ on, 145
- molecular friction, effect of, on, 108, 109
- prolongation of recovery
- relation between, and stimulus, 134, 135
- staircase effect, 122, 186
- stimulant, effect of, on, 141
- temperature, effect of, on, 111
- uniform, 102, 184
- Minchin on photo-electric cell, 165
- Molecular
- Munck on electric response in sensitive plants, 14
- Muscle, fatigue in, 38, 39, 40, 42. See also Fatigue
- Myograph, 2
- Negative variation, response by method of,
- Nerve,
- Nomenclature, anomalies of present, 9, 85
- Photographic recorder, 11, 22
- Plant
- chamber, 64
- electrical response
- radial E.M. response in, 49
- Poison,
- Record, simultaneous mechanical and electrical, of response, 13
- Recorder,
- Response-curve,
- characteristics of, 3
- electrical,
- abnormal,
- abolition of,
- additive effect of stimulus on,
- anæsthetics, effect of, on,
- annealing, effect of, on, in metal, 101, 138
- by method of block, 28, 82, 92
- by negative variation, 9, 18, 87, 183
- by relative depression, 87
- by relative exaltation, 89
- conditions for obtaining, 6, 86, 87
- continuous transformation from positive to negative in metal, 115
- decline and reversal of, under continuous light in photo-sensitive cell, 166
- decline and reversal of, under continuous light in retina, 166
- depressants, effect of, on inorganic, 142
- diminution of. See Fatigue
- diphasic
- dose, effect of,
- enhancement of, after continuous stimulation in metal, 127, 128, 186
- enhancement of, after continuous stimulation in nerve, 127, 186
- maximum effect due to superposition of stimulus, 35, 136
- measure of physiological activity, 13
- molecular
- physiological character of, in plant, 30
- positive and negative, 11
- prolongation of recovery in, by ‘poison’ in metal, 145
- prolongation of recovery in, by ‘poison’ in muscle, 144
- prolongation of recovery in, from overstrain, 106
- relation between, and stimulus
- staircase effect,
- stimulant, effect of, on, in metal, 141
- temperature, effect of, on. See Temperature
- threshold of, 135
- to light. See Light
- uniform
- universal applicability of, 12
- mechanical, 1
- retinal. See Light
- simultaneous mechanical and electrical record of, 13
- Retina. See Light
- Sanderson, Burdon-, on electrical response in sensitive plants, 14
- Spring-tapper, mechanical stimulus by, 23
- Staircase effect
- Steiner on retinal response, 149
- Stimuli, maximum effect due to superposition of,
- Stimulus,
- Waller
- on enhancement of nerve-response after continuous stimulation, 127
- on relation between stimulus and response in muscle, nerve, and retina, 52, 162
- on retinal response, 150, 156, 165
- on reversal of response in stale nerve and retina, 11, 124, 164
- on transformation from abnormal to normal response in nerve after continuous stimulation, 124
PRINTED BY
SPOTTISWOODE AND CO. LTD., NEW-STREET SQUARE
LONDON
PRINTED BY
SPOTTISWOODE AND CO. LTD., NEW STREET SQUARE
LONDON
Download ePUB
If you like this ebook, consider a donation!