This is a modern-English version of Sex-linked Inheritance in Drosophila, originally written by Morgan, Thomas Hunt, Bridges, Calvin B. (Calvin Blackman).
It has been thoroughly updated, including changes to sentence structure, words, spelling,
and grammar—to ensure clarity for contemporary readers, while preserving the original spirit and nuance. If
you click on a paragraph, you will see the original text that we modified, and you can toggle between the two versions.
Scroll to the bottom of this page and you will find a free ePUB download link for this book.
| Transcriber's note: |
A few typographical errors have been corrected. They
appear in the text like this, and the
explanation will appear when the mouse pointer is moved over the marked
passage. |
SEX-LINKED INHERITANCE IN
DROSOPHILA
BY
BY
T. H. MORGAN and C. B. BRIDGES
T. H. MORGAN and C. B. BRIDGES

WASHINGTON
Published by the Carnegie Institution of Washington
1916
WASHINGTON
Published by the Carnegie Institution of Washington
1916
CARNEGIE INSTITUTION OF WASHINGTON
Publication No. 237.
CARNEGIE INSTITUTION OF WASHINGTON
Pub No. 237.
PRESS OF GIBSON BROTHERS, INC.
WASHINGTON, D. C.
PRESS OF GIBSON BROTHERS, INC.
WASHINGTON, D.C.
CONTENTS.
| PAGE. | |
| Part I. Introduction | 5 |
| Mendel's law of segregation | 5 |
| Linkage and chromosomes | 5 |
| Crossing-over | 7 |
| The Y chromosome and non-disjunction | 8 |
| Mutation in Drosophila ampelophila | 10 |
| Multiple allelomorphs | 11 |
| Sex-linked lethals and the sex ratio | 14 |
| Influence of the environment on the realization of two sex-linked characters | 16 |
| Sexual polymorphism | 17 |
| Fertility and sterility in the mutants | 18 |
| Balanced inviability | 19 |
| How the factors are located in the chromosomes | 20 |
| The sex-linked factors of Drosophila | 21 |
| Map of chromosome X | 22 |
| Nomenclature | 24 |
| Part II. New info | 25 |
| White | 25 |
| Rudimentary | 25 |
| Miniature | 26 |
| Vermilion | 27 |
| Yellow | 27 |
| Abnormal abdomen | 27 |
| Eosin | 28 |
| Bifid | 28 |
| Linkage of bifid with yellow, with white, and with vermilion | 29 |
| Linkage of cherry, bifid, and vermilion | 30 |
| Reduplicated legs | 31 |
| Lethal 1 | 31 |
| Lethal 1a | 32 |
| Spot | 33 |
| Sable | 34 |
| Linkage of yellow and sable | 35 |
| Linkage of cherry and sable | 37 |
| Linkage of eosin, vermilion, and sable | 37 |
| Linkage of miniature and sable | 40 |
| Linkage of vermilion, sable, and bar | 40 |
| Dot | 44 |
| Linkage of vermilion and dot | 44 |
| Bow | 46 |
| Bow by arc | 47 |
| Lemon body-color | 48 |
| Linkage of cherry, lemon, and vermilion | 48 |
| Lethal 2 | 49 |
| Cherry | 51 |
| A system of quadruple allelomorphs | 51 |
| Linkage of cherry and vermilion | 51 |
| Compounds of cherry | 52 |
| Fused | 53 |
| Linkage of eosin and fused | 54 |
| Linkage of vermilion, bar, and fused | 56 |
| {4} Forked | 58 |
| Linkage of vermilion and forked | 59 |
| Linkage of cherry and forked | 59 |
| Linkage of forked, bar, and fused | 60 |
| Linkage of sable, rudimentary, and forked | 61 |
| Linkage of rudimentary, forked, and bar | 62 |
| Shifted | 63 |
| Linkage of shifted and vermilion | 63 |
| Linkage of shifted, vermilion, and bar | 64 |
| Lethals sa and sb | 64 |
| Bar | 66 |
| Notch | 66 |
| Depressed | 67 |
| Linkage of depressed and bar | 67 |
| Linkage of cherry, depressed, and vermilion | 68 |
| Club | 69 |
| Genotypic club | 70 |
| Linkage of club and vermilion | 70 |
| Linkage of yellow, club, and vermilion | 70 |
| Linkage of cherry, club, and vermilion | 72 |
| Green | 73 |
| Chrome | 74 |
| Lethal 3 | 74 |
| Lethal 3a | 75 |
| Lethal 1b | 76 |
| Facet | 76 |
| Linkage of facet, vermilion, and sable | 77 |
| Linkage of eosin, facet, and vermilion | 78 |
| Lethal sc | 79 |
| Lethal sd | 79 |
| Furrowed | 80 |
| Additional data for yellow, white, vermilion, and miniature | 80 |
| New data contributed by A. H. Sturtevant and H. J. Muller | 82 |
| Summary of the previously determined cross-over values | 83 |
| Summary of all data upon linkage of gens in chromosome I. | 84 |
| References. | 86 |
PART I. INTRODUCTORY.
MENDEL'S LAW OF SEGREGATION.
Mendel's Law of Segregation.
Although the ratio of 3 to 1 in which contrasted characters reappear in the second or F2 generation is sometimes referred to as Mendel's Law of Heredity, the really significant discovery of Mendel was not the 3 to 1 ratio, but the segregation of the characters (or rather, of the germinal representatives of the characters) which is the underlying cause of the appearance of the ratio. Mendel saw that the characters with which he worked must be represented in the germ-cells by specific producers (which we may call factors), and that in the fertilization of an individual showing one member of a pair of contrasting characters by an individual showing the other member, the factors for the two characters meet in the hybrid, and that when the hybrid forms germ-cells the factors segregate from each other without having been contaminated one by the other. In consequence, half the germ-cells contain one member of the pair and the other half the other member. When two such hybrid individuals are bred together the combinations of the pure germ-cells give three classes of offspring, namely, two hybrids to one of each of the pure forms. Since the hybrids usually can not be distinguished from one of the pure forms, the observed ratio is 3 of one kind (the dominant) to 1 of the other kind (the recessive).
Although the 3 to 1 ratio in which contrasting traits appear in the second or F2 generation is often called Mendel's Law of Heredity, Mendel's most important discovery wasn't the 3 to 1 ratio itself, but
There is another discovery that is generally included as a part of Mendel's Law. We may refer to this as the assortment in the germ-cells of the products of the segregation of two or more pairs of factors. If assortment takes place according to chance, then definite F2 ratios result, such as 9:3:3:1 (for two pairs) and 27:9:9:9:3:3:3:1 (for three pairs), etc. Mendel obtained such ratios in peas, and until quite recently it has been generally supposed that free assortment is the rule when several pairs of characters are involved. But, as we shall try to show, the emphasis that has been laid on these ratios has obscured the really important part of Mendel's discovery, namely, segregation; for with the discovery in 1906 of the fact of linkage the ratios based on free assortment were seen to hold only for combinations of certain pairs of characters, not for other combinations. But the principle of segregation still holds for each pair of characters. Hence segregation remains the cardinal point of Mendelism. Segregation is to-day Mendel's Law.
There's another finding that is usually considered part of Mendel's Law. We can call this the assortment in the germ cells resulting from the segregation of two or more pairs of factors. If assortment occurs by chance, then specific F2 ratios emerge, like 9:3:3:1 (for two pairs) and 27:9:9:9:3:3:3:1 (for three pairs), and so on. Mendel got these ratios in peas, and until fairly recently, it was widely believed that free assortment is the norm when multiple pairs of traits are involved. However, as we will try to demonstrate, the focus on these ratios has overshadowed the truly significant aspect of Mendel's discovery, which is segregation; because after the discovery in 1906 of linkage, it became clear that the ratios based on free assortment only apply to certain combinations of traits, not all of them. Still, the principle of segregation remains valid for each pair of traits. Therefore, segregation is the key concept of Mendelism. Segregation is what Mendel's Law is all about today.
LINKAGE AND CHROMOSOMES.
LINKS AND CHROMOSOMES.
It has been found that when certain characters enter a cross together (i. e., from the same parent) their factors tend to pass into the same gamete of the hybrid, with the result that other ratios than the chance ratios described by Mendel are found in the F2 generation. {6}Such cases of linkage have been described in several forms, but nowhere on so extensive a scale as in the pomace fly, Drosophila ampelophila. Here, over a hundred characters that have been investigated as to their linkage relations are found to fall into four groups, the members of each group being linked, in the sense that they tend to be transmitted to the gametes in the same combinations in which they entered from the parents. The members of each group give free assortment with the members of any of the other three groups. A most significant fact in regard to the linkage shown by the Drosophila mutants is that the number of linked groups corresponds to the number of pairs of the chromosomes. If the gens for the Mendelian characters are carried by the chromosomes we should expect to find demonstrated in Drosophila that there are as many groups of characters that are inherited together as there are pairs of chromosomes, provided the chromosomes retain their individuality. The evidence that the chromosomes are structural elements of the cell that perpetuate themselves at every division has continually grown stronger. That factors have the same distribution as the chromosomes is clearly seen in the case of sex-linked characters, where it can be shown that any character of this type appears in those individuals which from the known distribution of the X chromosomes must also contain the chromosome in question. For example, in Drosophila, as in many other insects, there are two X chromosomes in the cells of the female and one X chromosome in the cells of the male. There is in the male, in addition to the X, also a Y chromosome, which acts as its mate in synapsis and reduction. After reduction each egg carries an X chromosome. In the male there are two classes of sperm, one carrying the X chromosome and the other carrying the Y chromosome. Any egg fertilized by an X sperm produces a female; any egg fertilized by a Y sperm produces a male. The scheme of inheritance is as follows.
It has been found that when certain characters enter a cross together (i.e., from the same parent), their factors tend to pass into the same gamete of the hybrid, resulting in different ratios than the chance ratios described by Mendel in the F2 generation. {6} These cases of linkage have been described in several forms, but nowhere on such a large scale as in the pomace fly, Drosophila ampelophila. Here, over a hundred characters that have been studied for their linkage relations fall into four groups, with members of each group being linked in the sense that they tend to be transmitted to the gametes in the same combinations they entered from the parents. Members of each group assort freely with those of any of the other three groups. A very significant fact regarding the linkage shown by the Drosophila mutants is that the number of linked groups corresponds to the number of pairs of chromosomes. If the genes for the Mendelian characters are carried by the chromosomes, we would expect to find in Drosophila that there are as many groups of characters that are inherited together as there are pairs of chromosomes, assuming the chromosomes maintain their individuality. Evidence that chromosomes are structural elements of the cell that replicate themselves during each division has continually grown stronger. The distribution of factors matching that of chromosomes is clearly seen in the case of sex-linked characters, where it can be shown that any character of this type appears in individuals that, based on the known distribution of the X chromosomes, must also contain the chromosome in question. For example, in Drosophila, as in many other insects, females have two X chromosomes in their cells, while males have one X chromosome. Additionally, the male has a Y chromosome, which pairs with the X during synapsis and reduction. After reduction, each egg carries an X chromosome. In males, there are two types of sperm: one carrying the X chromosome and the other carrying the Y chromosome. An egg fertilized by an X sperm produces a female, while an egg fertilized by a Y sperm produces a male. The scheme of inheritance is as follows.
| Eggs | X—X |
| Sperm | X—Y |
| Daughter | XX |
| Son | XY |
The sons get their single X chromosome from their mother, and should therefore show any character whose gen is carried by such a chromosome. In sex-linked inheritance all sons show the characters of their mother. A male transmits his sex-linked character to his daughters, who show it if dominant and conceal it if recessive. But any daughter will transmit such a character, whether dominant or recessive, to half of her sons. The path of transmission of the gen is the same as the path followed by the X chromosome, received here {7}from the male. Many other combinations show the same relations. In the case of non-disjunction, to be given later, there is direct experimental evidence of such a nature that there can no longer be any doubt that the X chromosomes are the carriers of certain gens that we speak of as sex-linked. This term (sex-linked) is intended to mean that such characters are carried by the X chromosome. It has been objected that this use of the term implies a knowledge of a factor for sex in the X chromosome to which the other factors in that chromosome are linked; but in fact we have as much knowledge in regard to the occurrence of a sex factor or sex factors in the X chromosome as we have for other factors. It is true we do not know whether there is more than one sex-factor, because there is no crossing-over in the male (the heterozygous sex), and crossing-over in the female does not influence the distribution of sex, since like parts are simply interchanged. It follows from this that we are unable as yet to locate the sex factor or factors in the X chromosome. The fact that we can not detect crossing-over under this condition is not an argument against the occurrence of linkage. We are justified, therefore, in speaking of the factors carried by the X chromosome as sex-linked.
Sons receive their single X chromosome from their mother, which means they will display any traits associated with that chromosome. In sex-linked inheritance, all sons show the traits of their mother. A male passes his sex-linked traits to his daughters, who will express them if they're dominant and hide them if they're recessive. Any daughter will then pass on this trait, whether dominant or recessive, to half of her sons. The transmission path of the gene follows the same route as the X chromosome, which is inherited from the father. Many other combinations show similar patterns. In cases of non-disjunction, which will be discussed later, there is clear experimental evidence that the X chromosomes carry certain genes referred to as sex-linked. This term (sex-linked) means that such traits are associated with the X chromosome. Some have argued that this usage suggests a knowledge of a sex factor in the X chromosome that other factors are linked to; however, we know as much about the existence of sex factors in the X chromosome as we do about other factors. It is true we are unclear whether there is more than one sex factor, since males (the heterozygous sex) do not experience crossing-over, and crossing-over in females does not affect sex distribution because like parts are just swapped. Consequently, we cannot yet identify the sex factor or factors within the X chromosome. The inability to detect crossing-over under these conditions does not disprove the existence of linkage. Therefore, we can rightfully refer to the factors carried by the X chromosome as sex-linked.
CROSSING-OVER.
Crossover.
When two or more sex-linked factors are present in a male they are always transmitted together to his daughters, as must necessarily be the case if they are carried by the unpaired X chromosome. If such a male carrying, let us say, two sex-linked factors, is mated to a wild female, his daughters will have one X chromosome containing the factors for both characters, derived from the father, and another X chromosome that contains the factors that are normal for these two factors (the normal allelomorphs). The sons of such a female will get one or the other of these two kinds of chromosomes, and should be expected to be like the one or the other grandparent. In fact, most of the sons are of these two kinds. But, in addition, there are sons that show one only of the two original mutant characters. Clearly an interchange has taken place between the two X chromosomes in the female in such a way that a piece of one chromosome has been exchanged for the homologous piece of the other. The same conclusion is reached if the cross is made in such a way that the same two sex-linked characters enter, but, one from the mother and the other from the father. The daughter gets one of her sex chromosomes from her mother and the other from her father. She should produce, then, two kinds of sons, one like her mother and one like her father. In fact, the majority of her sons are of these two kinds, but, in addition, there are two other kinds of sons, one kind showing both mutant characters, the other kind showing normal characters. Here again the results must be due to interchange between the two X's in the hybrid female. The number of {8}the sons due to exchange in the two foregoing crosses is always the same, although they are of contrary classes. Clearly, then, the interchange takes place irrespective of the way in which the factors enter the cross. We call those classes that arise through interchange between the chromosomes "cross-over classes" or merely "cross-overs." The phenomenon of holding together we speak of as linkage.
When a male has two or more sex-linked traits, he always passes them together to his daughters, since they are carried by the unpaired X chromosome. If this male, for instance, carries two sex-linked traits and is paired with a wild female, his daughters will inherit one X chromosome from him that contains both traits and another X chromosome from their mother that has the normal versions of those traits. The sons of this female will receive either of these two types of chromosomes and will resemble one of their grandparents. In most cases, the sons will fall into these two categories. However, there are also some sons that exhibit only one of the two original mutant traits. This indicates that an exchange has occurred between the two X chromosomes in the female, resulting in a swap of segments. The same conclusion can be drawn when the mating happens with the same two sex-linked traits, but with one coming from the mother and the other from the father. The daughter receives one sex chromosome from her mother and the other from her father, so she should produce two types of sons: one like her mother and one like her father. Indeed, most of her sons belong to these two types, but there are also two other types, one showing both mutant traits and the other showing the normal traits. Again, this must result from an exchange between the two X chromosomes in the hybrid female. The number of {8}sons resulting from the exchange in the two previous crosses is always the same, even though they belong to different classes. Therefore, it’s clear that the exchange happens regardless of how the traits are inherited in the cross. We refer to the classes that emerge from this chromosome exchange as "cross-over classes" or simply "cross-overs." We refer to the phenomenon of traits being inherited together as linkage.
By taking a number of factors into consideration at the same time it has been shown that crossing-over involves large pieces of the chromosomes. The X chromosomes undergo crossing-over in about 60 per cent of the cases, and the crossing-over may occur at any point along the chromosome. When it occurs once, whole ends (or halves even) go over together and the exchange is always equivalent. If crossing-over occurs twice at the same time a middle piece of one chromosome is intercalated between the ends of the other chromosome. This process is called double crossing-over. It occurs not oftener than in about 10 per cent of cases for the total length of the X chromosome. Triple crossing-over in the X chromosome is extremely rare and has been observed only about a half dozen times.
By considering several factors at once, it has been shown that crossing-over involves large segments of the chromosomes. The X chromosomes experience crossing-over in about 60 percent of cases, and it can happen at any point along the chromosome. When it occurs once, entire ends (or even halves) switch places together, and the exchange is always equal. If crossing-over happens twice at the same time, a middle segment of one chromosome is inserted between the ends of the other chromosome. This process is known as double crossing-over. It occurs in no more than about 10 percent of cases across the total length of the X chromosome. Triple crossing-over in the X chromosome is extremely rare and has only been seen about half a dozen times.
While the genetic evidence forces one to accept crossing-over between the sex chromosomes in the female, that evidence gives no clue as to how such a process is brought about. There are, however, certain facts familiar to the cytologist that furnish a clue as to how such an interchange might take place. When the homologous chromosomes come together at synapsis it has been demonstrated, in some forms at least, that they twist about each other so that one chromosome comes to lie now on the one side now on the other of its partner. If at some points the chromosomes break and the pieces on the same side unite and pass to the same pole of the karyokinetic spindle, the necessary condition for crossing-over will have been fulfilled.
While the genetic evidence compels us to acknowledge crossing-over between the sex chromosomes in females, it doesn't provide any insight into how this process occurs. However, there are some facts known to cytologists that offer clues about how such an interchange might happen. When the homologous chromosomes align during synapsis, it has been shown, at least in some organisms, that they twist around each other. This causes one chromosome to occasionally move to one side and then to the other side of its partner. If the chromosomes break at certain points and the segments on the same side join together and migrate to the same pole of the karyokinetic spindle, then the essential condition for crossing-over will have been met.
THE Y CHROMOSOME AND NON-DISJUNCTION.
THE Y CHROMOSOME AND NON-DISJUNCTION.
Following Wilson's nomenclature, we speak of both X and Y as sex chromosomes. Both the cytological and the genetic evidence shows that when two X chromosomes are present a female is produced, when one, a male. This conclusion leaves the Y chromosome without any observed relation to sex-determination, despite the fact that the Y is normally present in every male and is confined to the male line. The question may be asked, and in fact has been asked, why may not the presence of the Y chromosome determine that a male develop and its absence that a female appear? The only answer that has yet been given, outside of the work on Drosophila, is that since in some insects there is no Y chromosome, there is no need to make such an assumption. But in Drosophila direct proof that Y has no such function is furnished by the evidence discovered by Bridges in the case of non-disjunction. (Bridges, 1913, 1914, 1916, and unpublished results.) {9}
Following Wilson's naming system, we refer to both X and Y as sex chromosomes. Both cytological and genetic evidence show that when there are two X chromosomes, a female is produced, and when there’s one, a male is produced. This conclusion leaves the Y chromosome seemingly unrelated to sex determination, even though the Y is typically found in every male and is only passed down through the male line. One might wonder, and has indeed wondered, why the presence of the Y chromosome doesn’t determine male development while its absence leads to female development. The only response given so far, aside from research on Drosophila, is that in some insects without a Y chromosome, there's no need to assume such a role for it. However, in Drosophila, direct evidence that Y does not have this function is provided by findings from Bridges regarding non-disjunction. (Bridges, 1913, 1914, 1916, and unpublished results.) {9}
Ordinarily all the sons and none of the daughters show the recessive sex-linked characters of the mother when the father carries the dominant allelomorph. The peculiarity of non-disjunction is that sometimes a female produces a daughter like herself or a son like the father, although the rest of the offspring are perfectly regular. For example, a vermilion female mated to a wild male produces vermilion sons and wild-type daughters, but rarely also a vermilion daughter or a wild-type son. The production of these exceptions (primary exceptions) by a normal XX female must be due to an aberrant reduction division at which the two X chromosomes fail to disjoin from each other. In consequence both remain in the egg or both pass into the polar body. In the latter case an egg without an X chromosome is produced. Such an egg fertilized by an X sperm produces a male with the constitution XO. These males received their single X from their father and therefore show the father's characters. While these XO males are exceptions to sex-linked inheritance, the characters that they do show are perfectly normal, that is, the miniature or the bar or other sex-linked characters that the XO male has are like those of an XY male, showing that the Y normally has no effect upon the development of these characters. But that the Y does play some positive rôle is proved by the fact that all the XO males have been found to be absolutely sterile.
Normally, all the sons and none of the daughters exhibit the recessive sex-linked traits of the mother when the father carries the dominant allele. The unusual circumstance of non-disjunction means that sometimes a female can have a daughter like herself or a son like the father, even though the rest of the offspring are completely typical. For example, a vermilion female paired with a wild male produces vermilion sons and wild-type daughters, but occasionally also a vermilion daughter or a wild-type son. The occurrence of these exceptions (primary exceptions) in a normal XX female must be due to an abnormal reduction division where the two X chromosomes do not separate. As a result, both stay in the egg or both go into the polar body. In the latter case, an egg without an X chromosome is formed. If this egg is fertilized by an X sperm, it results in a male with the constitution XO. These males receive their single X from their father and therefore display the father's traits. While these XO males are exceptions to sex-linked inheritance, the traits they exhibit are completely normal. In other words, the miniature, bar, or other sex-linked characteristics that the XO male has are similar to those of an XY male, indicating that the Y chromosome usually doesn't influence the development of these traits. However, the fact that all XO males are found to be completely sterile proves that the Y chromosome does play some positive role.
While the presence of the Y is necessary for the fertility of the male, it has no effect upon sex itself. This is shown even more strikingly by the phenomenon known as secondary non-disjunction. If the two X chromosomes that fail to disjoin remain in the egg, and this egg is fertilized by a Y sperm, an XXY individual results. This is a female which is like her mother in all sex-linked characters (a matroclinous exception), since she received both her X chromosomes from her mother and none from her father. As far as sex is concerned this is a perfectly normal female. The extra Y has no effect upon the appearance of the characters, even in the case of eosin, where the female is much darker than the male. The only effect which the extra Y has is as an extra wheel in the machinery of synapsis and reduction; for, on account of the presence of the Y, both X's of the XXY female are sometimes left within the ripe egg, a process called secondary non-disjunction. In consequence, an XXY female regularly produces exceptions (to the extent of about 4 per cent). A small percentage of reductions are of this XX-Y type; the majority are X-XY. The XY eggs, produced by the X-XY reductions, when fertilized by Y sperm, give XYY males, which show no influence of the extra Y except at synapsis and reduction. By mating an XXY female to an XYY male, XXYY females have been produced and these are perfectly normal in appearance. We may conclude from the fact that visibly indistinguishable males have been produced with the formulas XO, XY, and XYY, and {10}likewise females with the formulas XX, XXY, and XXYY, that the Y is without effect either on the sex or on the visible characters (other than fertility) of the individual.
While the presence of the Y chromosome is essential for male fertility, it doesn't influence the sex itself. This is further illustrated by the phenomenon known as secondary non-disjunction. If the two X chromosomes that fail to separate remain in the egg, and this egg is fertilized by a Y sperm, an XXY individual is produced. This individual is a female who shares all sex-linked traits with her mother (a matroclinous exception), as she inherited both X chromosomes from her mother and none from her father. In terms of sex, this is a completely normal female. The extra Y chromosome does not affect the visible traits, even in cases like eosin, where the female is much darker than the male. The only role the extra Y plays is as an additional component in the processes of synapsis and reduction. Because of the Y's presence, both X chromosomes in the XXY female are sometimes retained in the mature egg, a process known as secondary non-disjunction. As a result, an XXY female regularly produces exceptions (about 4 percent). A small percentage of reductions are of this XX-Y type; most are X-XY. The XY eggs produced by the X-XY reductions, when fertilized by Y sperm, create XYY males, who show no influence from the extra Y, except during synapsis and reduction. By mating an XXY female with an XYY male, XXYY females have been created, and these appear perfectly normal. We can conclude that visibly indistinguishable males have been produced with the formulas XO, XY, and XYY, and likewise females with the formulas XX, XXY, and XXYY, indicating that the Y chromosome has no effect on the sex or on the visible characteristics (aside from fertility) of the individual.
The evidence is equally positive that sex is quantitatively determined by the X chromosome—that two X's determine a female and one a male. For in the case of non-disjunction, a zero or a Y egg fertilized by an X sperm produces a male, while conversely an XX egg fertilized by a Y sperm produces a female. It is thus impossible to assume that the X sperms are normally female-producing because of something else than the X or that the Y sperm produce males for any other reason than that they normally fertilize X eggs. Both the X and the Y sperm have been shown to produce the sex opposite to that which they normally produce when they fertilize eggs that are normal in every respect, except that of their X chromosome content. These facts establish experimentally that sex is determined by the combinations of the X chromosomes, and that the male and female combinations are the causes of sex differentiation and are not simply the results of maleness and femaleness already determined by some other agent.
The evidence clearly shows that sex is primarily determined by the X chromosome—two X's create a female and one X with a Y creates a male. In cases of non-disjunction, a zero or Y egg fertilized by an X sperm results in a male, while an XX egg fertilized by a Y sperm results in a female. Therefore, it’s not possible to say that X sperms typically produce females for any reason other than the X chromosome, or that Y sperm create males for any reason other than the fact that they typically fertilize X eggs. Both X and Y sperm have been shown to produce the opposite sex when they fertilize normal eggs, except for the differences in X chromosome content. These facts experimentally confirm that sex is determined by the combinations of X chromosomes, and that the male and female combinations are the reasons for sex differentiation, rather than being simply effects of an already established maleness or femaleness from another source.
Cytological examination has demonstrated the existence of one XXYY female, and has checked up the occurrence in the proper classes and proportions of the XXY females. Numerous and extensive breeding-tests have been made upon the other points discussed. The evidence leaves no escape from the conclusion that the genetic exceptions are produced as a consequence of the exceptional distribution of the X chromosomes and that the gens for the sex-linked characters are carried by those chromosomes.
Cytological examination has shown the presence of one XXYY female and has confirmed the occurrence of XXY females in the right classes and proportions. Extensive breeding tests have been conducted on the other points discussed. The evidence makes it clear that the genetic exceptions arise from the unusual distribution of the X chromosomes, and that the genes for the sex-linked traits are carried by those chromosomes.
MUTATION IN DROSOPHILA AMPELOPHILA.
Mutation in Drosophila ampelophila.
The first mutants were found in the spring of 1910. Since then an ever-increasing series of new types has been appearing. An immense number of flies have come under the scrutiny of those who are working in the Zoological Laboratory of Columbia University, and the discovery of so many mutant types is undoubtedly due to this fact. But that mutation is more frequent in Drosophila ampelophila than in some of the other species of Drosophila seems not improbable from an extensive examination of other types. It is true a few mutants have been found in other Drosophilas, but relatively few as compared with the number in D. ampelophila. Whether ampelophila is more prone to mutate, or whether the conditions under which it is kept are such as to favor this process, we have no knowledge. Several attempts that we have made to produce mutations have led to no conclusive results.
The first mutants were discovered in the spring of 1910. Since then, a growing number of new types have been appearing. A huge number of flies have been examined by researchers at the Zoological Laboratory of Columbia University, and the discovery of so many mutant types is definitely linked to this effort. However, it doesn't seem unlikely that mutation occurs more frequently in Drosophila ampelophila than in some other Drosophila species, based on a thorough examination of other types. It's true that a few mutants have been found in other Drosophilas, but there's a relatively small number compared to those in D. ampelophila. We don't know whether ampelophila is more likely to mutate or if the conditions in which it's kept somehow encourage this process. Several attempts we've made to induce mutations have not yielded any conclusive results.
The mutants of Drosophila have been referred to by Baur as "mutations through loss," but inasmuch as they differ in no respect that we can discover from other mutants in domesticated animals and plants, there is no particular reason for putting them into this category unless {11}to imply that new characters have not appeared, or that those that have appeared must be due to loss in the sense of absence of something from the germ-plasm.
The mutants of Drosophila have been described by Baur as "mutations through loss," but since we can't find any differences compared to other mutants in domesticated animals and plants, there's really no reason to categorize them this way unless we want to suggest that new traits haven’t emerged, or that those that have come about are a result of the absence of something in the germ-plasm. {11}
In regard to the first point, several of the mutants are characterized by what seem to be additions. For example, the eye-color sepia is darker than the ordinary red. At least three new markings have been added to the thorax. A speck has appeared at the base of the wing, etc. These are recessive characters, it is true, but the character "streak," which consists of a dark band added to the thorax, is a dominant. If dominance is supposed to be a criterion as to "presence," then it should be pointed out that among the mutants of Drosophila a number of dominant types occur. But clearly we are not justified by these criteria in inferring anything whatever in regard to the nature of the change that takes place in the germ-plasm. Probably the only data which give a basis for attempting to decide the nature of the change in the germ-plasm are from cases where multiple allelomorphs are found. Several such cases are known to us, and two of these are found in the X chromosome group, namely, a quadruple system (white, eosin, cherry, red), and a triple system (yellow, spot, gray). In such cases each member acts as the allelomorph of any other member, and only two can occur in any one female, and only one in any male. If the normal allelomorph is thought of as the positive character, which one of the mutants is due to its loss or to its absence? If each is produced by a loss it must be a different loss that acts as an allelomorph to the other loss. This is obviously absurd unless a different idea from the one usually promulgated in regard to "absence" is held.
Regarding the first point, several of the mutants are characterized by what seem to be additions. For instance, the eye color sepia is darker than the usual red. At least three new markings have appeared on the thorax. A speck has shown up at the base of the wing, etc. These are recessive traits, it's true, but the trait "streak," which consists of a dark band added to the thorax, is dominant. If dominance is considered a criterion for "presence," then it should be noted that among the mutants of Drosophila, several dominant types exist. However, we cannot draw any conclusions about the nature of the changes occurring in the germ-plasm based on these criteria. Probably, the only data that provide a basis for determining the nature of the change in the germ-plasm are from cases where multiple allelomorphs are identified. Several such cases are known to us, with two found in the X chromosome group: a quadruple system (white, eosin, cherry, red) and a triple system (yellow, spot, gray). In these cases, each member acts as an allelomorph of any other member, and only two can occur in any one female, while only one can occur in any male. If the normal allelomorph is viewed as the positive trait, which of the mutants results from its loss or absence? If each is the result of a loss, it must be a different loss that acts as an allelomorph to the other loss. This is clearly absurd unless we adopt a different interpretation than the one usually put forward regarding "absence."
MULTIPLE ALLELOMORPHS.
MULTIPLE ALLELES.
It appears that Cuénot was the first to find a case (in mice) in which the results could be explained on the basis that more than two factors may stand in the relation of allelomorphs to each other. In other words, a given factor may become the partner of more than one other factor, although, in any one individual, no more than two factors stand in this relation. While it appears that his evidence as published was not demonstrative, and that, at the time he wrote, the possibility of such results being due to very close linkage could not have been appreciated as an alternative explanation, nevertheless it remains that Cuénot was right in his interpretation of his results and that the factors for yellow, gray, gray white-belly, and black in mice form a system of quadruple allelomorphs.
It seems that Cuénot was the first to discover a case (in mice) where the results could be understood by considering that more than two factors might relate to each other as allelomorphs. In other words, a specific factor can partner with more than one other factor, although, in any single individual, only two factors have this relationship. While it seems that his published evidence was not conclusive and that the potential for such results to be due to very close linkage may not have been recognized as an alternative explanation at the time he wrote, it still holds that Cuénot was correct in his interpretation of his findings and that the factors for yellow, gray, gray white-belly, and black in mice create a system of quadruple allelomorphs.
There are at least two such systems among the factors in the first chromosome in Drosophila. The first of these includes the factor for white eyes, that for eosin eyes, and that for cherry eyes, and of course that allelomorph of these factors present in the wild fly and which when present gives the red color. In this instance the normal {12}allelomorph dominates all the other three, but in mice the mutant factor for yellow dominates the wild or "normal" allelomorph.
There are at least two such systems among the factors in the first chromosome of Drosophila. The first includes the factor for white eyes, the factor for eosin eyes, and the factor for cherry eyes, along with the wild type allelomorph that produces red color. In this case, the normal {12}allelomorph dominates the other three, while in mice, the mutant factor for yellow dominates the wild or "normal" allelomorph.
The other system of multiple allelomorphs in the first chromosome is a triple system made up of yellow (body-color), spot (on abdomen), and their normal allelomorph—the factor in the normal fly that stands for "gray."
The other system of multiple alleles on the first chromosome is a triple system consisting of yellow (body color), spot (on the abdomen), and their normal allele—the factor in the normal fly that represents "gray."
In general it may be said that there are two principal ways in which it is possible to show that certain factors (more than two) are the allelomorphs of each other. First, if they are allelomorphs only two can exist in the same individual; and, in the case of sex-linked characters, while two may exist in the same female, only one can exist in the male, for he contains but one X chromosome. Second, all the allelomorphs should give the same percentages of crossing-over with each other factor in the same chromosome.
In general, it can be said that there are two main ways to demonstrate that certain factors (more than two) are allelomorphs of one another. First, if they are allelomorphs, only two can be present in the same individual; for sex-linked traits, while two can be present in the same female, only one can be present in the male, since he has only one X chromosome. Second, all the allelomorphs should show the same percentages of crossing-over with each other factor on the same chromosome.
It is a question of considerable theoretical importance whether these cases of multiple allelomorphs are only extreme cases of linkage or whether they form a system quite apart from linkage and in relation to normal allelomorphism. It may be worth while, therefore, to discuss this question more at length, especially because Drosophila is one of the best cases known for such a discussion.
It’s a question of significant theoretical importance whether these cases of multiple alleles are just extreme examples of linkage or if they represent a system separate from linkage and related to normal alleles. So, it may be useful to explore this question in more detail, especially since Drosophila is one of the best-known examples for such a discussion.
The factors in the first chromosome are linked to each other in various degrees. When they are as closely linked as yellow body-color and white eyes crossing-over takes place only once in a hundred times. If two factors were still nearer together it is thinkable that crossing-over might be such a rare occurrence that it would require an enormous number of individuals to demonstrate its occurrence. In such a case the factors might be said to be completely linked, yet each would be supposed to have its normal allelomorph in the homologous chromosome of the wild type. Imagine, then, a situation in which one of these two mutant factors (a) enters from one parent and the other mutant factor (b) from the other parent. The normal allelomorph of a may be called A. It enters the combination with b, while the normal allelomorph B of b enters the combination with a. Since b is completely linked to A and a to B, the result will be the same as though a and b were the allelomorphs of each other, for in the germ-cells of the hybrid aBAb the assortment will be into aB and Ab, which is the same as though a and b acted as segregating allelomorphs.
The factors in the first chromosome are connected to each other to varying degrees. When they are as closely connected as yellow body color and white eyes, crossing over happens only once in a hundred times. If two factors were even closer together, it’s possible that crossing over could be so rare that it would take a huge number of individuals to show it happening. In this case, the factors might be considered completely linked, but each would still have its normal allelomorph on the homologous chromosome of the wild type. Now, imagine a situation where one of these two mutant factors (a) comes from one parent and the other mutant factor (b) comes from the other parent. The normal allelomorph of a can be labeled A. It combines with b, while the normal allelomorph B of b combines with a. Since b is completely linked to A and a is linked to B, the outcome will be the same as if a and b were allelomorphs of each other. In the germ cells of the hybrid aBAb, the assortment will be aB and Ab, which is the same as if a and b were segregating allelomorphs.
There is no way from Mendelian data by which this difference between a true case of multiple allelomorphs and one of complete linkage (as just illustrated) can be determined. There is, however, a different line of attack which, in a case like that of Drosophila, will give an answer to this question. The answer is found in the way in which the mutant factors arise. This argument has been fully developed in the book entitled "The Mechanism of Mendelian Inheritance," and will therefore not be repeated here. It must suffice to say that if two mutant {13}types that behave as allelomorphs of each other arise separately from the wild form, one of them must have arisen as a double mutation of two factors so close to each other as to be completely linked—a highly improbable occurrence when the infrequency of mutations is taken into consideration.[1] The evidence opposed to such an interpretation is now so strong that there can be little doubt that multiple allelomorphs have actually appeared.
There’s no way to determine the difference between a true case of multiple alleles and one of complete linkage using Mendelian data, as demonstrated earlier. However, there’s another approach that, in cases like that of Drosophila, can provide an answer to this question. The answer is found in how the mutant factors occur. This argument has been thoroughly explained in the book titled "The Mechanism of Mendelian Inheritance," so I won’t repeat it here. It’s enough to say that if two mutant types that behave like alleles of each other arise separately from the wild type, one of them must have resulted from a double mutation of two factors that are so close together that they are completely linked—a very unlikely event when considering the rarity of mutations. The evidence against such an interpretation is now so strong that there is little doubt that multiple alleles have actually emerged.
On a priori grounds there is no reason why several mutative changes might not take place in the same locus of a chromosome. If we think of a chromosome as made up of a chain of chemical particles, there may be a number of possible recombinations or rearrangements within each particle. Any change might make a difference in the end-product of the activity of the cell, and give rise to a new mutant type. It is only when one arbitrarily supposes that the only possible change in a factor is its loss that any serious difficulty arises in the interpretation of multiple allelomorphs.
On a priori grounds, there’s no reason why several mutational changes can’t occur in the same spot on a chromosome. If we think of a chromosome as a chain of chemical particles, there could be various possible recombinations or rearrangements within each particle. Any change could impact the end result of the cell's activity and lead to a new mutant type. It’s only when we assume that the only possible change in a factor is its loss that any significant issues come up in understanding multiple allelomorphs.
One of the most striking facts connected with the subject of multiple allelomorphs is that the same kind of change is effected in the same organ. Thus, in the quadruple system mentioned above, the color of the eye is affected. In the yellow-spot system the color of the body is involved. In mice it is the coat-color that is different in each member of the series. While this is undoubtedly a striking relation and one which seems to fit well with the idea that such effects are due to mutative changes in the same fundamental element that affects the character in question, yet on the other hand it would be dangerous to lay too much emphasis on this point, because any given organ may be affected by other factors in a similar manner, and also because a factor frequently produces more than a single effect. For instance, the factor that when present gives a white eye affects also the general yellowish pigment of the body. If red-eyed and white-eyed flies are put for several hours into alcohol, the yellowish body-color of the white-eyed flies is freely extracted, but not that of the red-eyed flies. In the living condition the difference between the body-colors of the red- and of the white-eyed flies is too slight to be visible, but after extraction in alcohol the difference is striking. There are other effects also that follow in the wake of the white factor. Now, it is quite conceivable that in some specific case one of the effects might be more striking than the one produced in that organ more markedly affected by the other factor of the allelomorphic series. In such a case the relation mentioned above might seemingly disappear. For this reason it is well not to insist too strongly on the idea that multiple allelomorphs affect the same part in the same way, even although at present that appears to be the rule for all known cases.
One of the most striking facts associated with the topic of multiple alleles is that the same type of change occurs in the same organ. For example, in the quadruple system mentioned earlier, the eye color is affected. In the yellow-spot system, the body color is involved. In mice, it’s the coat color that varies among each member of the series. While this is undoubtedly a notable relationship and seems to align with the idea that such effects are due to mutations in the same fundamental element affecting the specific trait, it would be risky to place too much emphasis on this point. This is because any organ can be influenced by other factors in a similar way, and also because a single factor often produces multiple effects. For instance, the factor that causes a white eye also influences the general yellowish pigment of the body. If red-eyed and white-eyed flies are placed in alcohol for several hours, the yellowish body color of the white-eyed flies is easily extracted, but not that of the red-eyed flies. In their living state, the difference between the body colors of the red-eyed and white-eyed flies is too subtle to see, but after extraction in alcohol, the difference is noticeable. There are also other effects that occur as a result of the white factor. It’s entirely possible that in some specific cases, one of these effects might be more pronounced than the one produced in the organ that is more significantly impacted by the other factor in the allelic series. In such cases, the previously mentioned relationship might seem to vanish. Therefore, it’s wise not to insist too strongly on the idea that multiple alleles affect the same part in the same way, even though that currently appears to be the norm for all known cases.
SEX-LINKED LETHALS AND THE SEX RATIO.
SEX-LINKED LETHALS AND THE SEX RATIO.
Most of the mutant types of Drosophila show characteristics that may be regarded as superficial in so far as they do not prevent the animal from living in the protected life that our cultures afford. Were they thrown into open competition with wild forms, or, better said, were they left to shift for themselves under natural conditions, many or most of the types would no doubt soon die out. So far as we can see, there is no reason to suppose that the mutations which can be described as superficial are disproportionally more likely to occur than others. Of course, superficial mutations are more likely to survive and hence to be seen; while if mutations took place in important organs some of them would be expected to affect injuriously parts essential to the life of the individual and in consequence such an individual perishes. The "lethal factors" of Drosophila may be supposed to be mutations of some such nature; but as yet we have not studied this side of the question sufficiently, and this supposed method of action of the lethals is purely speculative. Whatever the nature of the lethals' action, it can be shown that from among the offspring obtained from certain stocks expected classes are missing, and the absence of these classes can be accounted for on the assumption that there are present mutant factors that follow the Mendelian rule of segregation and which show normal linkage to other factors, but whose only recognizable difference from the normal is the death of those individuals which receive them. The numerical results can be handled in precisely the same way as are other linkage results.
Most types of mutant Drosophila exhibit traits that might be seen as superficial since they don’t stop the organism from thriving in the sheltered environments our cultures provide. If they were subjected to direct competition with wild types, or better yet, if they had to fend for themselves in natural settings, many or most of these types would likely go extinct fairly quickly. From what we can observe, there's no reason to think that superficial mutations are significantly more likely to happen than others. Naturally, superficial mutations are more likely to survive and thus be detected; on the other hand, mutations affecting vital organs would probably harm essential parts for the individual’s survival, leading to their death. The so-called “lethal factors” in Drosophila might be mutations of this kind; however, we haven't explored this aspect thoroughly yet, and the proposed mechanism for the lethals is purely hypothetical. Regardless of how the lethals operate, it's clear that when we look at the offspring from certain stocks, expected categories are missing. This absence can be explained by the presence of mutant factors that adhere to Mendelian segregation rules and show normal linkage to other factors, with their only noticeable difference being the death of individuals that inherit them. The numerical results can be analyzed exactly like other linkage results.
There are some general relations that concern the lethals that may be mentioned here, while the details are left for the special part or are found in the special papers dealing with these lethals. A factor of this kind carried by the X chromosome would be transmitted in the female line because the female, having two X chromosomes, would have one of them with the normal allelomorph (dominant) of the lethal factor carried by the other X chromosome. Half of her sons would get one of her X's, the other half the other. Those sons that get the lethal X will die, since the male having only one X lacks the power of containing both the lethal and its normal allelomorph. The other half of the sons will survive, but will not transmit the lethal factor. In all lethal stocks there are only half as many sons as daughters. The heterozygous lethal-bearing female, fertilized by a normal male, will give rise to two kinds of daughters; one normal in both X's, the other with a normal X and a lethal-bearing X chromosome. The former are always normal in behavior, and the latter repeat in their descendants the 2:1 sex-ratio.
There are some general relationships regarding lethals that can be discussed here, while the specifics are reserved for the special section or can be found in the special papers addressing these lethals. A factor of this nature carried by the X chromosome would be passed down through the female line because females, having two X chromosomes, would possess one with the normal allele (dominant) for the lethal factor carried on the other X chromosome. Half of her sons will inherit one of her X's, while the other half will inherit the other. Those sons who inherit the lethal X will not survive, as males, having only one X, cannot carry both the lethal allele and its normal counterpart. The other half of the sons will survive but will not pass on the lethal factor. In all lethal stocks, there are only half as many sons as daughters. A heterozygous female carrying the lethal factor, when fertilized by a normal male, will produce two types of daughters: one with normal X chromosomes and the other with one normal X and one lethal-bearing X chromosome. The former will always be normal in behavior, while the latter will repeat the 2:1 sex ratio in their offspring.
Whether a female bearing the same lethal twice (i.e., one homozygous for a given lethal) would die, can not be stated, for no such females are obtainable, because the lethal males, which alone could bring about {15}such a condition, do not exist. The presumption is that a female of this kind would also die if the lethal acts injuriously on some vital function or structure.
Whether a female carrying the same lethal trait twice (i.e., one homozygous for a specific lethal) would die cannot be determined, as no such females are available because the lethal males, which could create this situation, do not exist. It is assumed that a female like this would also die if the lethal affects some vital function or structure negatively.
Since only half of the daughters of the lethal-bearing females carry the lethal, the stock can be maintained by breeding daughters separately in each generation to insure obtaining one which repeats the 2:1 ratio. There is, however, a much more advantageous way of carrying on the stock—one that also confirms the sufficiency of the theory.
Since only half of the daughters of the lethal-bearing females carry the lethal trait, the population can be sustained by breeding daughters separately in each generation to ensure that one maintains the 2:1 ratio. However, there is a much better way to manage the population—one that also supports the validity of the theory.
In carrying on a stock of a lethal, advantage can be taken of linkage. A lethal factor has a definite locus in the chromosome; if, then, a lethal-bearing female is crossed to a male of another stock with a recessive character whose factor lies in the X chromosome very close to the lethal factor, half the daughters will have lethal in one X and the recessive in the other. The lethal-bearing females can be picked out from their sisters by the fact that they give a 2:1 sex-ratio, and by the fact that nearly all the sons that do survive show the recessive character. If such females are tested by breeding to the recessive males, then the daughters which do not show the recessive carry the lethal, except in the few cases of crossing-over. Thus in each generation the normal females are crossed to the recessive males with the assurance that the lethal will not be lost. If instead of the single recessive used in this fashion, a double recessive of such a sort that one recessive lies on each side of the lethal is used, then in each generation the females which show neither recessive will almost invariably contain the lethal, since a double cross-over is required to remove the lethal.
In maintaining a stock of a lethal trait, you can take advantage of genetic linkage. A lethal factor is located at a specific spot on the chromosome; therefore, when a female carrying the lethal trait is crossed with a male from another stock that has a recessive trait located on the X chromosome close to the lethal factor, half of the daughters will carry the lethal trait on one X chromosome and the recessive trait on the other. The females that carry the lethal trait can be identified among their sisters because they produce a 2:1 sex ratio, and nearly all the surviving sons display the recessive trait. If these females are bred with recessive males, the daughters that do not show the recessive trait carry the lethal factor, except in rare cases of crossover. Thus, in each generation, normal females are bred with recessive males, ensuring that the lethal trait will not be lost. If, instead of using a single recessive trait in this manner, a double recessive trait is used—one recessive on each side of the lethal trait—then in each generation, the females that do not show either recessive trait will almost always carry the lethal factor, as a double crossover is needed to eliminate it.
It is true that females carrying two different lethals might arise and not die, because the injurious effect of each lethal would be dominated by its allelomorph in the other X chromosome. Such females can not be obtained by combining two existing lethals, since lethal males do not survive. They can occur only through a new lethal arising through mutation in the homologous chromosome of a female that already carries one lethal. Rare as such an event must be, it has occurred in our cultures thrice. The presence of a female of this kind will be at once noticed by the fact that she produces no sons, or very rarely one, giving in consequence extraordinary sex-ratios. The rare appearance of a son from such a female can be accounted for in the following way: If crossing-over occurs between her X chromosomes the result will be that one X will sometimes contain two lethals, the other none. The latter, if it passes into a male, will lead to the development of a normal individual. The number of such males depends on the distance apart of the two lethals in the chromosome. There is a crucial test of this hypothesis of two lethals in females giving extraordinary ratios. This test has been applied to the cases in which such females were found, by Rawls (1913), by Morgan (1914c), and again by Stark (1915), and it has been found to confirm the explanation. The daughters of {16}such a female should all (excepting a rare one due to crossing-over) give 2:1 ratios, because each daughter must get one or the other X chromosome of her mother, that is, one or the other lethal. Although the mother was fertilized by a normal male, every daughter is heterozygous for one or the other of the lethal factors. The daughters of the two-lethal females differ from the daughters of the one-lethal female in that the former mother, as just stated, gives all lethal-bearing daughters; the latter transmits her lethal to only half of her daughters.
It is true that females carrying two different lethals might arise and survive because the harmful effect of each lethal is overshadowed by its counterpart on the other X chromosome. Such females cannot be created by combining two existing lethals, since lethal males don’t survive. They can only occur through a new lethal arising from a mutation in the homologous chromosome of a female that already carries one lethal. As rare as this event is, it has happened in our cultures three times. The presence of such a female is immediately noticeable because she produces no sons, or very rarely one, resulting in unusual sex ratios. The rare appearance of a son from such a female can be explained as follows: If crossing-over occurs between her X chromosomes, one X may sometimes contain both lethals while the other contains none. If the latter X goes to a male, it will lead to the development of a normal individual. The number of such males depends on the distance between the two lethals on the chromosome. There is a crucial test for this hypothesis of two lethals in females producing unusual ratios. This test has been applied to cases where such females were found, by Rawls (1913), by Morgan (1914c), and again by Stark (1915), and it has confirmed the explanation. The daughters of {16}such a female should all (except for a rare one due to crossing-over) show 2:1 ratios, because each daughter must inherit one of her mother's X chromosomes, meaning one of the lethals. Although the mother was fertilized by a normal male, every daughter is heterozygous for one of the lethal factors. The daughters of the two-lethal females differ from those of the one-lethal female in that the former mother, as mentioned, produces all lethal-carrying daughters; the latter only passes her lethal to half of her daughters.
INFLUENCE OF THE ENVIRONMENT ON THE REALIZATION OF TWO SEX-LINKED CHARACTERS.
INFLUENCE OF THE ENVIRONMENT ON THE REALIZATION OF TWO SEX-LINKED CHARACTERS.
The need of a special environment in order that certain mutant characters may express themselves has been shown for abnormal abdomen (Morgan, 1912d, 1915b) and for reduplication of the legs (Hoge, 1915). In a third type, club, described here (page 69), the failure of the unfolding of the wing which occurs in about 20 per cent of the flies is also without much doubt an environmental effect, but as yet the particular influence that causes the change is unknown.
The necessity of a specific environment for certain mutant traits to manifest has been demonstrated for abnormal abdomens (Morgan, 1912d, 1915b) and for duplicated legs (Hoge, 1915). In a third type, known as club, described here (page 69), the failure of wing unfolding seen in about 20 percent of the flies is also likely an environmental effect, but the exact influence causing this change remains unknown.
A very extensive series of observations has been made on the character called abnormal abdomen. In pure cultures kept moist with abundance of fresh food all the flies that hatch for the first few days have the black bands of the abdomen obliterated or made faint and irregular. As the bottles get dry and the food becomes scarce the flies become more and more normal, until at last they are indistinguishable from the normal flies. Nevertheless these normal-looking flies will give rise in a suitable environment to the same kind of flies as the very abnormal flies first hatched. By breeding from the last flies of each culture, and in dry cultures, flies can be bred from normal ancestors for several generations, and then by making the conditions favorable for the appearance of the abnormal condition, the flies will be as abnormal as though their ancestors had always been abnormal. Here, then, is a character that is susceptible to the variations in the environment, yet whatever the realized condition of the soma may be, that condition has no effect whatever on the nature of the germ-plasm. A more striking disproof of the theory of the inheritance of acquired characters would be hard to find.
A very extensive series of observations has been made on a condition known as abnormal abdomen. In pure cultures maintained in moist conditions with plenty of fresh food, all the flies that hatch during the first few days have their black abdominal bands either removed or appear faint and irregular. As the bottles dry out and food becomes limited, the flies start to look more and more normal, until finally they become indistinguishable from regular flies. However, these normal-looking flies can still produce the same type of flies as the very abnormal ones that hatched initially, given the right conditions. By breeding from the last flies of each culture and using dry cultures, flies can be produced from normal ancestors for several generations, and then by creating favorable conditions for the abnormal condition to arise, the flies will appear as abnormal as if their ancestors had always been abnormal. This demonstrates a characteristic that is influenced by environmental changes, yet whatever the current state of the body may be, that state has no effect on the nature of the germ-plasm. A more compelling counterexample to the theory of the inheritance of acquired traits would be hard to find.
A demonstration is given in this instance of the interaction between a given genotypic constitution and a special environment. The character abnormal is a sex-linked dominant. Therefore, if an abnormal male is mated to a wild female the daughters are heterozygous for abnormal, while the sons, getting their X chromosome from their mother, are entirely normal. In a wet environment all the daughters are abnormal and the sons normal. As the culture dries out the daughters' color becomes normal in appearance. But while the sons {17}will never transmit abnormality to any of their descendants in any environment, the daughters will transmit (if bred to normal males) in a suitable environment their peculiarity to half of their daughters and to half of their sons. The experiment shows convincingly that the abnormal abdomen appears in a special environment only in those flies that have a given genotypic constitution.
A demonstration is provided here of how a specific genetic makeup interacts with a particular environment. The abnormal trait is a sex-linked dominant. So, when an abnormal male mates with a wild female, the daughters are heterozygous for the abnormal trait, while the sons, receiving their X chromosome from their mother, are completely normal. In a wet environment, all the daughters show the abnormal trait, and the sons are normal. As the environment dries, the daughters' color starts to look normal. However, while the sons {17} will never pass on the abnormal trait to any of their offspring regardless of the environment, the daughters will pass it on (if bred with normal males) in a suitable environment to half of their daughters and half of their sons. This experiment clearly demonstrates that the abnormal abdomen only appears in a specific environment among flies that have a particular genetic constitution.
As the cultures dry out the abnormal males are the first to change over to normal, then the heterozygous females, and lastly the homozygous females. It is doubtful if any far-reaching conclusion can be drawn from this series, because the first and second classes differ from each other not only in the presence of one or of two factors for abnormal, but also by the absence in the first case (male) of an entire X chromosome with its contained factors. The second and third classes differ from each other only by the abnormal factor.
As the cultures dry out, the abnormal males change to normal first, followed by the heterozygous females, and lastly the homozygous females. It’s unlikely that any significant conclusions can be made from this sequence because the first and second groups are different not only because one has one factor for abnormality and the other has two, but also because the males lack an entire X chromosome with its associated factors. The second and third groups differ only by the abnormal factor.
Similar results were found in the mutant type called reduplicated legs, which is a sex-linked recessive character that appears best when the cultures are kept at about 10° C. As Miss M. A. Hoge has shown, this character then becomes realized in nearly all of the flies that have the proper constitution, but not in flies of normal constitution placed in the same environment. Here the effect is produced by cold.
Similar results were observed in the mutant type known as reduplicated legs, which is a sex-linked recessive trait that shows best when the cultures are kept at around 10° C. As Miss M. A. Hoge demonstrated, this trait appears in almost all of the flies with the right genetic makeup, but not in normally constituted flies placed in the same conditions. In this case, the effect is caused by the cold.
SEXUAL POLYMORPHISM.
Sexual Dimorphism.
Outside the primary and secondary sexual differences between the male and the female, there is a considerable number of species of animals with more than one kind of female or male. Darwin and his followers have tried to explain such cases on the grounds that more than one kind of female (or male) might arise through natural selection, in consequence of some individuals mimicking a protected species. It is needless to point out here how involved and intricate such a process would be, because the mutation theory has cut the Gordian knot and given a simpler solution of the origin of such diandromorphic and digynomorphic conditions.
Outside the basic differences between male and female, there are many species of animals that have more than one type of female or male. Darwin and his followers have tried to explain these situations by suggesting that multiple types of females (or males) could arise through natural selection, as some individuals mimic a protected species. It's unnecessary to elaborate on how complex and complicated this process would be, since the mutation theory has offered a straightforward solution to the origin of these diandromorphic and digynomorphic conditions.
In Drosophila a mutant, eosin eye-color, appeared in which the female has darker eyes than the male. If such stock is crossed with cherry (another sex-linked recessive mutant, allelomorphic to eosin) the females in the F2 generation are alike (for the pure eosin and the eosin-cherry compound are not separable), but the cherry males and the eosin males are quite different in appearance. Here we have a simulation, at least, of a diandromorphic species. Such a group perpetuates itself, giving one type of female (inasmuch as eosin and cherry females are very closely similar) and two types of males, only one of which is like the females. A population of this kind is very directly comparable to certain polymorphic types that occur in nature. In Colias philodice there is one type of male, yellow, and two types of females, yellow and {18}white. In Colias eurydice the male is orange and the females are orange or white. In Papilio turnus the male is yellow and the females either yellow or black. Those cases are directly comparable to an eosin-cherry population, except that in Lepidoptera the female is heterozygous for the sex differential, in Diptera the male.
In Drosophila, a mutant called eosin eye-color appeared, where females have darker eyes than males. If this stock is crossed with cherry (another sex-linked recessive mutant, allelic to eosin), the females in the F2 generation are the same (since the pure eosin and the eosin-cherry combination can't be separated), but the cherry males and eosin males look quite different. This simulates a diandromorphic species. Such a group reproduces itself, yielding one type of female (given that eosin and cherry females are very similar) and two types of males, only one of which resembles the females. A population like this is very similar to certain polymorphic types found in nature. In Colias philodice, there is one type of male, yellow, and two types of females, yellow and white. In Colias eurydice, the male is orange, and the females are either orange or white. In Papilio turnus, the male is yellow, and the females can be yellow or black. These cases are directly comparable to an eosin-cherry population, except that in Lepidoptera, the female is heterozygous for the sex differential, while in Diptera, the male is.
Since in Drosophila the results are explicable on a sex-linked basis, a similar explanation may apply to polymorphism in butterflies. By suitable combinations of eosin and cherry most of the cases of polymorphism in butterflies may be simulated. To simulate the more complex cases, such as that of Papilio polytes and memnon, another allelomorph like eosin would have to be introduced. A population of mixed cherry and white would give three somatic types of females (cherry, cherry-white, and white) and two of males (cherry and white).
Since in Drosophila the results can be explained based on sex-linked traits, a similar explanation might apply to the color variations in butterflies. By using the right mixtures of eosin and cherry, most instances of color variations in butterflies can be replicated. To mimic the more complex cases, like those of Papilio polytes and memnon, another allele similar to eosin would need to be added. A population with a mix of cherry and white colors would result in three types of female butterflies (cherry, cherry-white, and white) and two types of males (cherry and white).
FERTILITY AND STERILITY IN THE MUTANTS.
FERTILITY AND STERILITY IN THE MUTANTS.
Aside from the decrease in fertility that occurs in certain stocks (a question that need not be treated here), there are among the types described in the text two cases that call for special comment. When the mutant type called "rudimentary" was first discovered, it was found that the females were sterile but the males were fully fertile. Later work has revealed the nature of the sterility of the female. The ovaries are present and in the young flies appear normal, but while in the normal flies the eggs in the posterior portion enlarge rapidly during the first few days after hatching, in the rudimentary females only a very few (about 15) eggs enlarge. The other eggs in the ovary remain at a lower stage of their development. Rarely the female lays a few eggs; when she does so some of the eggs hatch, and if she has been mated to a rudimentary male, the offspring are rudimentary females and males. The rudimentary females mate in the normal time with rudimentary or with normal males, and their sexual behavior is normal. Their sterility is therefore due to the failure of the eggs to develop properly. Whether in addition to this there is some incompatibility between the sperm and the eggs of this type (as supposed to be the case at one time) is not conclusively disproved, but is not probable from the evidence now available.
Aside from the drop in fertility that happens in certain stocks (a topic we won't discuss here), there are two cases among the types described in the text that need special attention. When the "rudimentary" mutant type was first discovered, it was observed that the females were sterile while the males were completely fertile. Further research has clarified the cause of the female sterility. The ovaries are present and look normal in young flies, but while normal flies see their eggs in the back grow rapidly in the first few days after hatching, rudimentary females only have a small number (about 15) of eggs that grow. The other eggs in the ovary stay at an earlier stage of development. Occasionally, the female lays a few eggs; when she does, some of those eggs hatch, and if she mated with a rudimentary male, the offspring are rudimentary females and males. The rudimentary females mate normally with either rudimentary or normal males, and their sexual behavior is typical. Their sterility is thus due to the eggs not developing properly. Whether there’s also some incompatibility between the sperm and the eggs of this type (as was once thought) hasn’t been definitively disproved, but current evidence makes it seem unlikely.
In the mutant called "fused" the females are sterile both with wild males and with males from their own stock. An examination of the ovaries of these females, made by Mr. C. McEwen, shows clearly that there are fewer than the normal number of mature eggs, recalling the case of rudimentary.
In the mutant known as "fused," the females are sterile both with wild males and with males from their own line. An examination of the ovaries of these females, conducted by Mr. C. McEwen, clearly shows that there are fewer than the normal number of mature eggs, similar to the case of rudimentary.
It should be noticed that there is no apparent relation between the sterility of these two types and the occurrence of the mutation in the X chromosome, because other mutations in the X do not cause sterility, and there is sterility in other mutant types that are due to factors in other chromosomes. {19}
BALANCED INVIABILITY.
BALANCED NONVIABILITY.
The determination of the cross-over values of the factors was at first hindered because of the poor viability of some of the mutants. If the viability of each mutant type could be determined in relation to the viability of the normal, "coefficients of viability" could serve as corrections in working with the various mutant characters. But it was found (Bridges and Sturtevant, 1914) that viability was so erratic that coefficients might mislead. At the same time it was becoming more apparent that poor viability is no necessary attribute of a character, but depends very largely on the condition of culture. Competition among larvæ was found to be the chief factor in viability. Mass cultures almost invariably have extremely poor viability, even though an attempt is made to supply an abundance of food. Special tests (Morgan and Tice, 1914) showed that even those mutants which were considered the very poorest in viability were produced in proportions fairly close to the theoretical when only one female was used for each large culture bottle and the amount and quality of food was carefully adjusted.
The determination of the crossover values for the factors was initially hindered by the poor viability of some of the mutants. If the viability of each mutant type could be assessed in relation to the viability of the normal type, "coefficients of viability" could be used as adjustments when working with different mutant traits. However, it was found (Bridges and Sturtevant, 1914) that viability was so unpredictable that these coefficients could be misleading. At the same time, it became clearer that poor viability is not an inherent trait of a character, but largely depends on the culture conditions. Competition among larvae was identified as the main factor affecting viability. Mass cultures consistently showed extremely low viability, even with efforts to provide ample food. Specific tests (Morgan and Tice, 1914) revealed that even those mutants thought to have the worst viability were produced in proportions fairly close to the theoretical when only one female was used for each large culture bottle and the quantity and quality of food was carefully managed.
For the majority of mutants which did well even under heavy competition in mass cultures the pair-breeding method reduced the disturbances due to viability to a point where they were negligible.
For most mutants that performed well even in tough competition in mass cultures, the pair-breeding method minimized the issues caused by viability to a level where they were barely noticeable.
Later a method was devised (Bridges, 1915) whereby mutations of poor viability could be worked with in linkage experiments fairly accurately and whereby the residual inviability of the ordinary characters could be largely canceled. This method consists in balancing the data of a certain class with poor viability by means of an equivalent amount of data in which the same class occurs as the other member of the ratio. Thus in obtaining data upon any linkage case it is best to have the total number of individuals made up of approximately equal numbers derived from each of the possible ways in which the experiment may be conducted. In the simplest case, in which the results are of the form AB:Ab:aB:ab, let us suppose that the class ab has a disproportionately low viability. If, then, ab occurs in an experiment as a cross-over class, that class will be too small and a false linkage value will be calculated. The remedy is to balance the preceding data by an equal amount of data in which ab occurs as a non-cross-over. In these latter the error will be the opposite of the previous one, and by combining the two experiments the errors should be balanced to give a better approximation to the true value. When equal amounts of data, secured in these two ways, are combined, all four classes will be balanced in the required manner by occurring both as non-cross-overs and as cross-overs. The error, therefore, should be very small. For three pairs of gens there are eight classes, and in order that each of them may appear as a non-cross-over, as each single cross-over, and as the double cross-over, four experiments must be made. {20}
Later, a method was developed (Bridges, 1915) that allowed mutations with low viability to be accurately used in linkage experiments while mostly canceling out the remaining inviability of regular traits. This method involves balancing the data from a specific class with low viability by using an equal amount of data where the same class appears as the other part of the ratio. Therefore, when gathering data for any linkage case, it’s best to ensure that the total number of individuals consists of roughly equal amounts from all the possible ways the experiment can be conducted. In the simplest scenario, where the results are in the form AB:Ab:aB:ab, let’s say that the class ab has significantly low viability. If ab shows up in an experiment as a cross-over class, that class will be too small, leading to an inaccurate linkage value. The solution is to balance the earlier data with an equivalent amount of data where ab appears as a non-cross-over. In these latter cases, the error will be opposite to the previous one, and by combining the results from both experiments, the errors should offset, giving a closer approximation to the true value. When equal amounts of data from these two methods are combined, all four classes will be balanced as they appear both as non-cross-overs and as cross-overs. Therefore, the error should be minimal. For three pairs of genes, there are eight classes, and to ensure each one appears as a non-cross-over, as a single cross-over, and as a double cross-over, four experiments must be conducted. {20}
HOW THE FACTORS ARE LOCATED IN THE CHROMOSOMES.
HOW THE FACTORS ARE LOCATED IN THE CHROMOSOMES.
A character is in the first chromosome if it is transmitted by the grandfather to half of his grandsons, while, in the reciprocal cross, the mother transmits her character to all her sons (criss-cross inheritance) and to half of her granddaughters and to half of her grandsons; in other words, if the factor that differentiates the character has the same distribution as the X chromosome. If, however, a new mutant type does not show this sex-linked inheritance, its chromosome is determined by taking advantage of the fact that in Drosophila there is no crossing-over in the male between factors in the same chromosome. For instance, if a new mutant type is found not to be sex-linked, its group is determined by the following tests: It is crossed to black, whose factor is known to be in the second chromosome, and to pink, whose factor lies in the third chromosome. If the factor of the new form should happen to be in the second chromosome, then, in the cross with black, no double recessive can appear, so that the F2 proportion is 2:1:1:0; but with pink, the mutant type should give the proportion 9:3:3:1, typical of free assortment.
A trait is in the first chromosome if it's passed down from the grandfather to half of his grandsons, while in the reciprocal cross, the mother passes her trait to all her sons (criss-cross inheritance) and to half of her granddaughters and half of her grandsons. In other words, if the factor that defines the trait has the same distribution as the X chromosome. However, if a new mutant type doesn’t show this sex-linked inheritance, its chromosome is identified by taking advantage of the fact that in Drosophila, there is no crossing-over in males between factors on the same chromosome. For example, if a new mutant type is found not to be sex-linked, its group is identified through the following tests: It is crossed with black, whose factor is known to be on the second chromosome, and with pink, whose factor is on the third chromosome. If the factor of the new type happens to be on the second chromosome, then in the cross with black, no double recessive will appear, resulting in an F2 ratio of 2:1:1:0; with pink, the mutant type should yield a ratio of 9:3:3:1, which is typical of free assortment.
If, however, the factor of the new form is in the third chromosome, then, when crossed to black, the double recessive and the 9:3:3:1 proportion appear in F2. But when crossed to pink no double recessive appears in F2, and the proportion 2:1:1:0 occurs.
If the factor for the new trait is on the third chromosome, then when it's crossed with black, the double recessive and the 9:3:3:1 ratio show up in F2. But when it's crossed with pink, no double recessive appears in F2, and the ratio is 2:1:1:0.
If these tests show that the new mutant does not belong to either the second or third chromosome, that is, if both with black and with pink the 9:3:3:1 ratio is obtained, then by exclusion the factor lies in the fourth chromosome, in which as yet only two factors have been found.
If these tests indicate that the new mutant isn’t associated with either the second or third chromosome, meaning that both the black and pink variants show a 9:3:3:1 ratio, then by process of elimination, the factor must be located in the fourth chromosome, where only two factors have been identified so far.
We propose to give in a series of papers an account of the mutant races of Drosophila and the linkage shown in their inheritance. In this paper we shall consider only the members of the first chromosome, describing a large number of new mutants with their linkage relations and summarizing to date all the linkage data relating to the first chromosome. In later papers we propose to consider the members of the second, third, and fourth chromosomes.
We plan to present a series of papers discussing the mutant strains of Drosophila and how they are linked in inheritance. In this paper, we will focus solely on the first chromosome, detailing a significant number of new mutants along with their linkage relationships, and summarizing all the linkage data related to the first chromosome up to now. In future papers, we aim to examine the members of the second, third, and fourth chromosomes.
The list at the top of page 21 gives the names of the factors dealt with in this paper. They stand in the order of their discovery, the mutant forms reported here for the first time being starred.
The list at the top of page 21 shows the names of the factors discussed in this paper. They are arranged in the order they were discovered, with the mutant forms mentioned here for the first time marked with an asterisk.
In each experiment the percentage of crossing-over is found by dividing the number of the cross-overs by the sum of the non-cross-overs and the cross-overs, and multiplying this quotient by 100. The resulting percentages, or cross-over values, are used as measures of the distances between loci. Thus if the experiments give a cross-over value of 5 per cent for white and bifid, we say that white and bifid lie 5 units apart in the X chromosome. Other experiments show that yellow and white are about 1 unit apart, and that yellow and bifid are about 6 units apart. We can therefore construct a diagram with yellow as {21}the zero, with white at 1, and with bifid at 6. If we know the cross-over values given by a new mutant with any two mutants of the same chromosome whose positions are already determined, then we can locate the new factor with accuracy, and be able to predict the cross-over value which the new factor will give with any other factor whose position is plotted.
In each experiment, the percentage of crossing-over is calculated by dividing the number of cross-overs by the total of non-cross-overs and cross-overs, then multiplying that result by 100. The percentages obtained, or cross-over values, are used to measure the distances between loci. So, if the experiments show a cross-over value of 5 percent for white and bifid, we say that white and bifid are 5 units apart on the X chromosome. Other experiments indicate that yellow and white are about 1 unit apart, while yellow and bifid are roughly 6 units apart. We can then create a diagram with yellow as {21}the zero point, white at 1, and bifid at 6. If we know the cross-over values provided by a new mutant with any two mutants of the same chromosome whose positions are already identified, we can accurately locate the new factor and predict the cross-over value it will have with any other factor whose position is already mapped.
The sex-linked factors of Drosophila.
The sex-linked traits of fruit flies.
| Gen. | Part affected. | Figure. | Symbol. | Locus. | Date found. | Found by. | |
| White | Eye-color | 11 | w | 1.1 | May | 1910 | Morgan. |
| Rudimentary | Wings | A | r | 55.1 | June | 1910 | Morgan. |
| Miniature | Wings | 7-8 | m | 36.1 | Aug. | 1910 | Morgan. |
| Vermilion | Eye-color | 10 | v | 33.0 | Nov. | 1910 | Morgan. |
| Yellow | Body-color | 5 | y | 0.0 | Jan. | 1911 | Wallace. |
| Abnormal | Abdomen | 4 | A′ | 2.4 | July | 1911 | Morgan. |
| Eosin | Eye-color | 7-8 | we | 1.1 | Aug. | 1911 | Morgan. |
| Bifid | Wings | B | bi | 6.3 | Nov. | 1911 | Morgan. |
| Reduplicated | Legs | 34.7 | Nov. | 1911 | Hoge. | ||
| Lethal 1 | Life | l1 | 0.7 | Feb. | 1912 | Rawls. | |
| Lethal 1a* | Life | l1a | 3.3 | Mar. | 1912 | Rawls. | |
| Spot* | Body-color | 14-17 | ys | 0.0 | April | 1912 | Cattell. |
| Sable* | Body-color | 2 | s | 43.0 | July | 1912 | Bridges. |
| Dot* | Thorax | 33 ± | July | 1912 | Bridges. | ||
| Bow* | Wings | C | Aug. | 1912 | Bridges. | ||
| Lemon* | Body-color | 3 | lm | 17.5 | Aug. | 1912 | Wallace. |
| Lethal 2 | Life | l2 | 12.5± | Sept. | 1912 | Morgan. | |
| Cherry | Eye-color | 9 | wc | 1.1 | Oct. | 1912 | Safir. |
| Fused* | Venation | D | fu | 59.5 | Nov. | 1912 | Bridges. |
| Forked* | Bristles | E | f | 56.5 | Nov. | 1912 | Bridges. |
| Shifted* | Venation | F | sh | 17.8 | Jan. | 1913 | Bridges. |
| Lethal sa | Life | lsa | 23.7 | Jan. | 1913 | Stark. | |
| Bar | Eye-shape | 12-13 | B′ | 57.0 | Feb. | 1913 | Tice. |
| Notch | Wing | N′ | 2.6 | Mar. | 1913 | Dexter. | |
| Depressed* | Wing | G | dp | 18.0 | April | 1913 | Bridges. |
| Lethal sb | Life | lsb | 16.7 | April | 1913 | Stark. | |
| Club* | Wings | H | cl | 14.6 | May | 1913 | Morgan. |
| Green* | Body-color | May | 1913 | Bridges. | |||
| Chrome* | Body-color | Sept. | 1913 | Bridges. | |||
| Lethal 3 | Life | l3 | 26.5 | Dec. | 1913 | Morgan. | |
| Lethal 3a | Life | l3a | 19.5 | Jan. | 1914 | Morgan. | |
| Lethal 1b* | Life | l1b | 1.1- | Feb. | 1914 | Morgan. | |
| Facet* | Eye | fa | 2.2 | Feb. | 1914 | Bridges. | |
| Lethal sc | Life | lsc | 66.2 | April | 1914 | Stark. | |
| Lethal sd | Life | lsd | May | 1914 | Stark. | ||
| Furrowed | Eye | fw | 38.0 | Nov. | 1914 | Duncan. | |
The factors are located preferably by short distances (i.e., by those cases in which the amount of crossing-over is small), because when the amount of crossing-over is large a correction must be made for double crossing-over, and the correction can be best found through breaking up the long distances into short ones, by using intermediate points.
The factors are ideally located by short distances (i.e., in situations where the amount of crossing-over is minimal), because when the amount of crossing-over is high, a correction needs to be made for double crossing-over. This correction is most effectively determined by dividing the long distances into shorter ones using intermediate points.

Diagram I.
Diagram I.
Diagram I has been constructed upon the basis of all the data summarized in table 65 (p. 84) for the first or X chromosome. It shows the relative positions of the gens of the sex-linked characters of Drosophila. One unit of distance corresponds to 1 per cent of crossing-over. Since all distances are corrected for double crossing-over and for coincidence, the values represent the total of crossing-over between the loci. The uncorrected value obtained in any experiment with two loci widely separated will be smaller than the value given in the map.
Diagram I is based on all the data summarized in table 65 (p. 84) for the first or X chromosome. It displays the relative positions of the genes for sex-linked traits of Drosophila. One unit of distance equals 1 percent of crossing-over. Since all distances have been adjusted for double crossing-over and coincidence, the values represent the total crossing-over between the loci. The unadjusted value obtained from any experiment with two widely separated loci will be smaller than the value shown on the map.
It may be asked what will happen when two factors whose loci are more than 50 units apart in the same chromosome are used in the same experiment? One might expect to get more than 50 per cent of cross-overs with such an experiment, but double crossing-over becomes disproportionately greater the longer the distance involved, so that in experiments the observed percentage of crossing-over does not rise above 50 per cent. For example, if eosin is tested against bar, somewhat under 50 per cent of cross-overs are obtained, but if the distance of bar from eosin is found by summation of the component distances the interval for eosin bar is 56 units.
It might be asked what happens when two factors that are more than 50 units apart on the same chromosome are used in the same experiment. One might expect to see over 50 percent crossovers with such an experiment, but double crossing-over increases disproportionately with the distance involved, so in experiments, the observed percentage of crossing-over does not exceed 50 percent. For instance, if eosin is tested against bar, slightly less than 50 percent of crossovers are obtained, but if the distance from bar to eosin is determined by adding the component distances, the interval for eosin bar is 56 units.
In calculating the loci of the first chromosome, a system of weighting was used which allowed each case to influence the positions of the loci in proportion to the amount of the data. In this way advantage was taken of the entire mass of data.
In calculating the locations of the first chromosome, a weighting system was used that allowed each case to impact the positions of the locations based on the amount of data. This way, the entire dataset was utilized effectively.
The factors (lethal 1, white, facet, abnormal, notch, and bifid) which lie close to yellow were the first to be calculated and plotted. The next step was to determine very accurately the position of vermilion with respect to yellow. There are many separate experiments which influence this calculation and all were proportionately weighted. Then, using vermilion as the fixed point the factors (dot, reduplicated, miniature, and sable) which lie close to vermilion were plotted. The same process was repeated in locating bar with respect to vermilion and the factors about bar with reference to bar. The last step was to interpolate the factors (club, lethal 2, lemon, depressed, and shifted), which form a group about midway between yellow and vermilion. Of these, club is the only one whose location is accurate. The apparent closeness of the grouping of these loci is not to be taken as significant, for they have been placed only with reference to the distant points yellow and vermilion and not with respect to each other; furthermore, the data available in the cases of lemon and depressed are very meager.
The factors (lethal 1, white, facet, abnormal, notch, and bifid) that are close to yellow were the first to be calculated and plotted. The next step was to accurately determine the position of vermilion in relation to yellow. There are several individual experiments that affect this calculation, and all were weighted appropriately. Then, using vermilion as the reference point, the factors (dot, reduplicated, miniature, and sable) that are near vermilion were plotted. The same process was repeated to locate bar in relation to vermilion and the factors around bar with respect to bar. The final step was to interpolate the factors (club, lethal 2, lemon, depressed, and shifted), which form a group about halfway between yellow and vermilion. Among these, club is the only one accurately positioned. The apparent closeness of these groupings should not be considered significant, as they have only been placed in relation to the distant points yellow and vermilion, not in relation to each other; furthermore, the available data for lemon and depressed are quite limited.
The factors which are most important and are most accurately located are yellow, white (eosin), bifid, club, vermilion, miniature, sable, forked, and bar. Of these again, white (eosin), vermilion, and bar are of prime importance and will probably continue to claim first rank. Of the three allelomorphs, white, eosin, and cherry, eosin is the most useful. {24}
The most important factors that are clearly defined are yellow, white (eosin), bifid, club, vermilion, miniature, sable, forked, and bar. Among these, white (eosin), vermilion, and bar hold the highest significance and will likely remain at the top. Among the three allelomorphs—white, eosin, and cherry—eosin is the most valuable. {24}
NOMENCLATURE.
Naming conventions.
The system of symbols used in the diagrams and table headings is as follows: The factor or gen for a recessive mutant character is represented by a lower-case letter, as v for vermilion and m for miniature. The symbols for the dominant mutant characters bar, abnormal, and notch are B′, A′, and N′. There are now so many characters that it is impossible to represent all of them by a single letter. We therefore add a subletter in such cases, as bifid (bi), fused (fu), and lethal 2 (l2). In the case of multiple allelomorphs we usually use as the base of the symbol the symbol of that member of the system which was first found and add a letter as an exponent to indicate the particular member, as ys for spot, we for eosin, and wc for cherry. The normal allelomorphs of the mutant gens are indicated by the converse letter, as V for not-vermilion, Bi for not-bifid, and b′ for not-bar. In the table headings the normal allelomorphs are indicated by position alone without the use of a symbol.
The system of symbols used in the diagrams and table headings is as follows: The factor or gene for a recessive mutant trait is shown by a lowercase letter, like v for vermilion and m for miniature. The symbols for the dominant mutant traits bar, abnormal, and notch are B′, A′, and N′. There are so many traits now that it's impossible to represent all of them with just one letter. So, we add a subletter in those cases, like bifid (bi), fused (fu), and lethal 2 (l2). When it comes to multiple allelomorphs, we usually base the symbol on the first discovered member of the system and add a letter as an exponent to indicate the specific member, such as ys for spot, we for eosin, and wc for cherry. The normal allelomorphs of the mutant genes are indicated by the opposing letter, like V for not-vermilion, Bi for not-bifid, and b′ for not-bar. In the table headings, the normal allelomorphs are shown by their position alone without using a symbol.
Thus the symbol  indicates that the female in question carried eosin,
not-vermilion, and bar in one chromosome and not-eosin, vermilion, and
not-bar in the other. The symbol
indicates that the female in question carried eosin,
not-vermilion, and bar in one chromosome and not-eosin, vermilion, and
not-bar in the other. The symbol 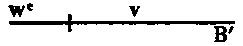 when used in the heading of a column in a table indicates that the
flies classified under this heading are the result of single
crossing-over between eosin and vermilion in a mother which was the
composition ; the symbol tells
at the same time that the flies that result from a single cross-over
between eosin and vermilion in the mother are of the two contrary
classes, eosin vermilion and bar. When a fly shows two or more
non-allelomorphic characters the names are written from left to right in
the order of their positions from the zero end of the map.
when used in the heading of a column in a table indicates that the
flies classified under this heading are the result of single
crossing-over between eosin and vermilion in a mother which was the
composition ; the symbol tells
at the same time that the flies that result from a single cross-over
between eosin and vermilion in the mother are of the two contrary
classes, eosin vermilion and bar. When a fly shows two or more
non-allelomorphic characters the names are written from left to right in
the order of their positions from the zero end of the map.
Thus the symbol indicates that the female in question carried eosin, not-vermilion, and bar on one chromosome and not-eosin, vermilion, and not-bar on the other. The symbol when used as a column header in a table indicates that the flies listed under this heading resulted from a single crossing-over between eosin and vermilion in a mother with the composition ; the symbol also indicates that the flies resulting from a single cross-over between eosin and vermilion in the mother belong to two opposite classes, eosin red and bar. When a fly shows two or more non-allelic traits, the names are written from left to right in the order of their positions from the zero end of the map.
PART II. NEW DATA.
WHITE.
WHITE.
(Plate II, figure 11.)
(__A_TAG_PLACEHOLDER_0__, figure 11.)
The recessive character white eye-color, which appeared in May 1910, was the first sex-linked mutation in Drosophila (Morgan, 1910a, 1910b). Soon afterwards (June 1910) rudimentary appeared, and the two types were crossed (Morgan, 1910c). Under the conditions of culture the viability of rudimentary was extremely poor, but the data demonstrated the occurrence of recombination of the factors in the ovogenesis so that white and rudimentary, though both sex-linked, were brought together into the same individual. The results were not fully recognized as linkage, because white and rudimentary are so far apart in the chromosome that they seemed to assort freely from each other.
The white eye-color trait, which is recessive, showed up in May 1910 and was the first sex-linked mutation in Drosophila (Morgan, 1910a, 1910b). Not long after, in June 1910, the rudimentary trait appeared, and the two types were crossed (Morgan, 1910c). Under the culture conditions, the survival rate of rudimentary was very low, but the data showed that recombination of the factors occurred during ovogenesis, allowing both white and rudimentary traits, even though they are sex-linked, to be present in the same individual. The findings were not completely understood as linkage because white and rudimentary are located far apart on the chromosome, making them appear to assort independently.
Owing to the excellent viability and the perfect sharpness of separation, white was extensively used in linkage experiments, especially with miniature and yellow (Morgan, 1911a; Morgan and Cattell, 1912 and 1913). White has been more extensively used than any other character in Drosophila, though it is now being used very little because of the fact that the double recessives of white with other sex-linked eye-colors, such as vermilion, are white, and consequently a separation into the true genetic classes is impossible. The place of white has been taken by eosin, which is an allelomorph of white and which can be readily used with any other eye-color.
Due to its great viability and the clear separation of traits, the white phenotype was widely used in linkage experiments, especially with miniature and yellow variations (Morgan, 1911a; Morgan and Cattell, 1912 and 1913). White has been used more than any other trait in Drosophila, but it's not used much anymore because the double recessives of white with other sex-linked eye colors, like vermilion, are also white. This makes it impossible to separate them into true genetic classes. Eosin, an allelomorph of white, has taken its place and can be easily used with any other eye color.
The locus of white and its allelomorphs is only 1.1 units from that of yellow, which is the zero of the chromosome. Yellow and white are very closely linked, therefore giving only about one cross-over per 100 flies.
The location of white and its variants is only 1.1 units away from yellow, which is at the start of the chromosome. Yellow and white are tightly linked, resulting in about one crossover per 100 flies.
All the published data upon the linkage of white with other sex-linked characters have been collected into table 65.
All the published data on the connection between white and other sex-linked traits have been compiled into table 65.
RUDIMENTARY.
BASIC.
Rudimentary, which appeared in June 1910, was the second sex-linked character in Drosophila (Morgan, 1910c). Its viability has always been very poor; in this respect it is one of the very poorest of the sex-linked characters. The early linkage data (Morgan, 1911a) derived from mass cultures have all been discarded. By breeding from a single F1 female in each large culture bottle it has been possible to obtain results which are fairly trustworthy (Morgan, 1912g; Morgan and Tice, 1914). These data appear in table 65, which summarizes all the published data. {26}
Rudimentary, which came out in June 1910, was the second sex-linked trait identified in Drosophila (Morgan, 1910c). Its viability has always been very low; in this regard, it is one of the least viable of the sex-linked traits. The initial linkage data (Morgan, 1911a) obtained from mass cultures has all been thrown out. By breeding from a single F1 female in each large culture bottle, it has been possible to get results that are fairly reliable (Morgan, 1912g; Morgan and Tice, 1914). This data is shown in table 65, which summarizes all the published information. {26}
The locus of rudimentary is at 55.1, for a long time the extreme right end of the known chromosome, though recently several mutants have been found to lie somewhat beyond it.
The basic location is at 55.1, which has long been the far right end of the known chromosome, although recently several mutants have been discovered to extend a bit beyond it.
 Fig. A. a. rudimentary wing; b. the wild fly for
comparison.
Fig. A. a. rudimentary wing; b. the wild fly for
comparison.
The rudimentary males are perfectly fertile, but the rudimentary females rarely produce any offspring at all, and then only a very few. The reason for this is that most of the germ-cells cease their development in the early growth stage of the eggs (Morgan, 1915a).
The basic males are completely fertile, but the basic females hardly ever produce any offspring, and when they do, it’s only a few. The reason for this is that most of the germ cells stop developing in the early growth stage of the eggs (Morgan, 1915a).
MINIATURE.
Mini.
(Plate II. figures 7 and 8.)
(__A_TAG_PLACEHOLDER_0__. figures 7 and 8.)
The recessive sex-linked mutant miniature wings appeared in August 1910 (Morgan, 1911b and 1912a). The viability of miniature is fair, and this stock has been used in linkage experiments more than any {27}other, with the single exception of white. While the wings of miniature usually extend backwards, they are sometimes held out at right angles to the body, and especially in acid bottles the miniature flies easily become stuck to the food or the wings become stringy, so that other wing characters are not easy to distinguish in those flies which are also miniature. At present vermilion, whose locus is at 33, in being used more frequently in linkage work. The locus of miniature at 36.1 is slightly beyond the middle of the chromosome.
The recessive sex-linked mutant miniature wings showed up in August 1910 (Morgan, 1911b and 1912a). The viability of miniature is decent, and this stock has been used in linkage experiments more than any {27}other, except for white. While the wings of miniature usually extend backward, they are sometimes held out at right angles to the body, and especially in acid bottles, the miniature flies can easily get stuck to the food or their wings become stringy, making it hard to distinguish other wing traits in those flies that are also miniature. Right now, vermilion, which is located at 33, is being used more often in linkage work. The locus of miniature at 36.1 is just slightly past the middle of the chromosome.
VERMILION.
Red.
(Plate II. figure 10.)
(__A_TAG_PLACEHOLDER_0__. figure 10.)
The recessive sex-linked mutant vermilion eye-color (Morgan, 1911c and 1912a) appeared in November 1910, and has appeared at least twice since then (Morgan and Plough, 1915). This is one of the best of the sex-linked characters, on account of its excellent viability, its sharp distinction from normal with very little variability, its value as a double recessive in combination with other sex-linked eye-colors, and because of its location at 33.0, very near to the middle of the known chromosome.
The recessive sex-linked mutant vermilion eye color (Morgan, 1911c and 1912a) showed up in November 1910 and has been seen at least twice since then (Morgan and Plough, 1915). This is one of the best sex-linked traits due to its strong viability, clear distinction from normal with very little variation, its usefulness as a double recessive when combined with other sex-linked eye colors, and its position at 33.0, very close to the center of the known chromosome.
YELLOW.
YELLOW.
(Plate I. figure 5.)
(__A_TAG_PLACEHOLDER_0__. figure 5.)
The recessive sex-linked mutant yellow body and wing-color appeared in January 1911 (Morgan, 1911c and 1912a). Its first appearance was in black stock; hence the fly was a double recessive, then called brown. Later the same mutation has appeared independently from gray stock. Yellow was found to be at the end of the X chromosome, and this end was arbitrarily chosen as the zero or the "left end," while the other gens are spoken of as lying at various distances to the right of yellow. Recently a lethal gen has been located less than one-tenth of a unit (-0.04) to the left of yellow, but yellow is still retained as the zero-point.
The recessive sex-linked mutant yellow body and wing color appeared in January 1911 (Morgan, 1911c and 1912a). Its first appearance was in black stock; so the fly was a double recessive, which was then called brown. Later, the same mutation appeared independently from gray stock. Yellow was found to be at the end of the X chromosome, and this end was arbitrarily designated as the zero or "left end," while the other genes are described as located at various distances to the right of yellow. Recently, a lethal gene was found less than one-tenth of a unit (-0.04) to the left of yellow, but yellow is still considered the zero point.
The viability of yellow is fairly good and the character can be separated from gray with great facility, and in consequence yellow has been used extensively, although at present it is being used less than formerly, since eosin lies only 1.1 units distant from yellow and is generally preferred.
The effectiveness of yellow is quite good, and it can be easily distinguished from gray. Because of this, yellow has been used a lot, although it’s currently being used less than before since eosin is just 1.1 units away from yellow and is generally favored.
ABNORMAL ABDOMEN.
ABNORMAL ABDOMEN.
(Plate I. figure 4.)
(__A_TAG_PLACEHOLDER_0__. fig. 4.)
The dominant sex-linked character abnormal abdomen appeared in July 1911 (Morgan, 1911d). It was soon found that the realization of the abnormal condition depended greatly upon the nature of the environment (Morgan, 1912). Recently a very extensive study of this character has been published (Morgan, 1915). As this case has been reviewed in the introduction, there is little further to be said here. {28}Because of the change that takes place as the culture grows older (the abnormal changing to normal), this character is not of much value in linkage work. The location of the factor in the X chromosome at 2.4 has been made out from the data given by Morgan (1915b). These data, which in general include only the abnormal classes, are summarized in table 1.
The main sex-linked trait of an abnormal abdomen showed up in July 1911 (Morgan, 1911d). It was quickly discovered that the expression of this abnormal condition relied heavily on the type of environment (Morgan, 1912). Recently, a comprehensive study of this trait was published (Morgan, 1915). Since this case has already been discussed in the introduction, there’s not much more to add here. {28}Due to the changes that occur as the culture ages (the abnormal turning normal), this trait isn't very useful for linkage studies. The position of the factor on the X chromosome at 2.4 has been determined from the data provided by Morgan (1915b). This data, which primarily includes only the abnormal classes, is summarized in table 1.
Table 1.—Linkage data, from Morgan, 1915b.
Table 1.—Linkage data, from Morgan, 1915b.
| Gens. | Total. | Cross- overs. | Cross-over values. |
| Yellow white | 28,018 | 334 | 1.2 |
| Yellow abnormal | 15,314 | 299 | 2.0 |
| White abnormal | 16,300 | 277 | 1.7 |
EOSIN.
EOSIN.
(Plate II, figures 7 and 8.)
(__A_TAG_PLACEHOLDER_0__, figures 7 and 8.)
The recessive sex-linked mutation eosin eye-color appeared in August 1911 in a culture of white-eyed flies (Morgan 1912a). The eye-color is different in the male and female, the male being a light pinkish yellow, while the female is a rather dark yellowish pink. Eosin is allelomorphic to white and the white-eosin compound or heterozygote has the color of the eosin male. There is probably no special significance in this coincidence of color, since similar dilutions to various degrees have been demonstrated for all the other eye-colors tested (Morgan and Bridges, 1913). Since eosin is allelomorphic to white, its locus is also at 1.1. Eosin is the most useful character among all those in the left end of the chromosome.
The recessive sex-linked mutation eosin eye color was first observed in August 1911 in a culture of white-eyed flies (Morgan 1912a). The eye color differs between males and females: males have a light pinkish yellow, while females have a darker yellowish pink. Eosin is allelic to white, and the white-eosin heterozygote displays the color of the eosin male. This color coincidence probably doesn't have any special significance since similar dilutions of various degrees have been shown for all the other eye colors that were tested (Morgan and Bridges, 1913). Given that eosin is allelic to white, its locus is also at 1.1. Eosin is the most useful characteristic among all those at the left end of the chromosome.
BIFID.
Bifid.
The sex-linked wing mutant bifid, which appeared in November 1911, is characterized by the fusion of all the longitudinal veins into a heavy stalk at the base of the wing. The wing stands out from the body at a wide angle, so that the fusion is easily seen. At the tip of the wing the third longitudinal vein spreads out into a delta which reaches to the marginal vein. The fourth longitudinal vein reaches the margin only rarely. There is very often opposite this vein a great bay in the margin, or the whole wing is irregularly truncated.
The sex-linked wing mutant bifid, first noted in November 1911, is defined by the merging of all the longitudinal veins into a thick stalk at the base of the wing. The wing juts out from the body at a wide angle, making the fusion clearly visible. At the tip of the wing, the third longitudinal vein expands into a delta shape that connects to the marginal vein. The fourth longitudinal vein rarely reaches the margin. Often, there is a large indentation in the margin opposite this vein, or the entire wing is oddly cut off.
The stock of bifid was at first extremely varied in the amount of this truncation. By selection a stock was secured which showed only very greatly reduced wings like those shown in figures a, b. Another stock (figs. c, d) was secured by outcrossing and selection which showed wings of nearly normal size and shape, which always had the bifid stalk, generally the spread positions (not as extreme), and often the delta and the shortened fourth longitudinal vein. We believe that the extreme reduction in size seen in the one stock was due to an added modifier of {29}the nature of beaded, since this could be eliminated by outcrossing and selection.
The bifid stock initially had a wide range in the extent of this truncation. Through selective breeding, a stock was developed that displayed only greatly reduced wings, like those in figures a and b. Another stock (figs. c and d) was created through outcrossing and selection, showing wings that were nearly normal in size and shape, always featuring the bifid stalk, typically in spread positions (though not as extreme), and often the delta along with a shortened fourth longitudinal vein. We think that the significant reduction in size observed in one stock was due to an additional modifier of {29}the beaded nature, which could be removed through outcrossing and selection.

Fig. B.—Bifid wing. c and d show the typical condition of bifid wings. All the longitudinal veins are fused into a heavy stalk at the base of the wing. a shows the typical position in which the bifid wings are held. The small size of the wings in a and b is due to the action of a modifier of the nature of "beaded" which has been eliminated in c, d.
Fig. B.—Bifid wing. c and d show the typical features of bifid wings. All the longitudinal veins are joined into a thick base at the bottom of the wing. a shows the common way bifid wings are held. The small size of the wings in a and b is due to a modifier with a "beaded" characteristic that has been removed in c and d.
LINKAGE OF BIFID WITH YELLOW, WITH WHITE, AND WITH VERMILION.
LINKAGE OF BIFID WITH YELLOW, WITH WHITE, AND WITH VERMILION.
The stock of the normal (not-beaded) bifid was used by Dr. R. Chambers, Jr., for determining the chromosome locus of bifid by means of its linkage relations to vermilion, white, and yellow (Chambers, 1913). We have attempted to bring together in table 2 the complete data and to calculate the locus of bifid.
The normal (not-beaded) bifid stock was used by Dr. R. Chambers, Jr., to find the chromosome location of bifid by analyzing its linkage relationships with vermilion, white, and yellow (Chambers, 1913). We have tried to compile all the data in table 2 and calculate the location of bifid.
Table 2.—Linkage data, from Chambers, 1913.
Table 2.—Linkage data, from Chambers, 1913.
| Gens. | Total. | Cross- overs. | Cross-over values. |
| Yellow bifid | 3,175 | 182 | 5.8 |
| White bifid | 20,800 | 1,127 | 5.3 |
| Bifid vermilion | 2,509 | 806 | 32.1 |
In the crosses between white and bifid there were 1,127 cross-overs in a total of 20,800 available individuals, which gives a cross-over value of 5.3. In the crosses between yellow and bifid there were 182 cross-overs in a total of 3,175 available individuals, which gives a cross-over value of 5.8. In crosses between bifid and vermilion there were 806 cross-overs in a total of 2,509, which gives a cross-over value of 32.1. On the basis of all the data summarized in table 65, bifid is located at 6.3 to the right of yellow.
In the crosses between white and bifid, there were 1,127 cross-overs out of 20,800 available individuals, resulting in a cross-over value of 5.3. In the crosses between yellow and bifid, there were 182 cross-overs out of 3,175 available individuals, giving a cross-over value of 5.8. For crosses between bifid and vermilion, there were 806 cross-overs out of 2,509, leading to a cross-over value of 32.1. Based on all the data summarized in table 65, bifid is positioned at 6.3 to the right of yellow.
LINKAGE OF CHERRY, BIFID, AND VERMILION.
LINKAGE OF CHERRY, BIFID, AND VERMILION.
In a small experiment of our own, three factors were involved—cherry, bifid, and vermilion. A cherry vermilion female was crossed to a bifid male. Two daughters were back-crossed singly to white bifid males. The female offspring will then give data for the linkage of cherry white with bifid, while the sons will show the linkage of the three gens, cherry, bifid, and vermilion. The results are shown in table 3.
In a small experiment we conducted, three factors were involved—cherry, bifid, and vermilion. A cherry vermilion female was crossed with a bifid male. Two daughters were back-crossed individually with white bifid males. The female offspring will then provide data on the linkage of cherry white with bifid, while the male offspring will demonstrate the linkage of the three genes: cherry, bifid, and vermilion. The results are shown in table 3.
Table 3.—P1 cherry vermilion ♀ ♀ × bifid ♂ ♂. B. C.[2] F1 wild-type ♀ × white bifid ♂ ♂.
Table 3.—P1 cherry vermilion female × bifid male. B. C.[2] F1 wild-type female × white bifid male.
| Refer- ence. | F2 females. | F2 males. | ||||||||||
| Non-cross- overs. | Cross-overs. | 
| 
| 
|  | |||||||
| White- cherry | Bifid. | White- cherry bifid. | Wild- type. | Cherry ver- milion. | Bifid. | Cherry bifid. | Ver- milion. | Cherry. | Bifid ver- milion. | Cherry bifid vermilion. | Wild- type. | |
| 262 | 40 | 46 | 1 | 2 | 45 | 38 | 3 | 2 | 11 | 13 | .. | .. |
| 263 | 47 | 45 | 3 | 3 | 30 | 50 | 1 | 3 | 8 | 10 | 1 | .. |
| Total. | 87 | 91 | 4 | 5 | 75 | 88 | 4 | 5 | 19 | 23 | 1 | 0 |
Both males and females give a cross-over value of 5 units for cherry bifid, which is the value determined by Chambers. The order of the factors, viz, cherry, bifid, vermilion, is established by taking advantage of the double cross-over classes in the males. The male classes give a cross-over value of 20 for bifid vermilion and 24 for cherry vermilion, which are low compared with values given by other experiments. The locus of bifid at 6.3 is convenient for many linkage problems, but this advantage is largely offset by the liability of the bifid flies to become stuck in the food and against the sides of the bottle. Bifid flies can be separated from the normal with certainty and with great ease. {31}
Both males and females have a crossover value of 5 units for cherry bifid, which is the value set by Chambers. The order of the factors—cherry, bifid, vermilion—is established by utilizing the double crossover classes in males. The male classes show a crossover value of 20 for bifid vermilion and 24 for cherry vermilion, which are low compared to the values from other experiments. The position of bifid at 6.3 is useful for many linkage problems, but this benefit is mostly countered by the tendency of bifid flies to get stuck in the food and against the sides of the bottle. Bifid flies can be easily and reliably separated from the normal ones. {31}
REDUPLICATED LEGS.
Duplicate legs.
In November 1912 Miss Mildred Hoge found that a certain stock was giving some males whose legs were reduplicated, either completely or only with respect to the terminal segments (described and figured, Hoge, 1915). Subsequent work by Miss Hoge showed that the condition was due to a sex-linked gen, but that at room temperature not all the flies that were genetically reduplicated showed reduplication. However, if the flies were raised through the pupa stage in the ice-box at a temperature of about 10° to 12° a majority of the flies which were expected to show reduplication did so. The most extremely reduplicated individual showed parts of 14 legs.
In November 1912, Miss Mildred Hoge discovered that a particular stock was producing some male flies with duplicated legs, either fully or just the end segments (as described and illustrated in Hoge, 1915). Further research by Miss Hoge revealed that this condition was caused by a sex-linked gene, but at room temperature, not all the genetically duplicated flies displayed reduplication. However, if the flies were raised through the pupal stage in the refrigerator at a temperature of about 10° to 12°, most of the flies that were expected to show reduplication did indeed do so. The most drastically reduplicated individual had parts of 14 legs.
In studying the cross-over values of reduplicated, only those flies that have abnormal legs are to be used in calculation, as in the case of abnormal abdomen where the phenotypically normal individuals are partly genetically abnormal. Table 4 gives a summary of the data secured by Miss Hoge.
In studying the cross-over values of reduplicated traits, only the flies with abnormal legs should be used in the calculations, just like in the case of abnormal abdomens where the phenotypically normal individuals are partly genetically abnormal. Table 4 provides a summary of the data collected by Miss Hoge.
Table 4.—Summary of linkage data upon reduplicated legs, from Hoge, 1915.
Table 4.—Summary of linkage data on reduplicated legs, from Hoge, 1915.
| Gens. | Total. | Cross- overs. | Cross-over values. |
| White reduplicated | 418 | 121 | 29.0 |
| Reduplicated vermilion | 667 | 11 | 1.7 |
| Reduplicated bar | 583 | 120 | 20.6 |
The most accurate data, those upon the value for reduplicated and vermilion, give for reduplicated a distance of 1.7 from vermilion, either to the right or to the left. The distance from white is 29, which would place the locus for reduplication to the left of vermilion, which is at 33. The data for bar give a distance of 21, but since bar is itself 24 units from vermilion, this distance of 21 would seem to place the locus to the right of vermilion. The evidence is slightly in favor of this position to the right of vermilion at 34.7, where reduplicated may be located provisionally. In any case the locus is so near to that of vermilion that final decision must come from data involving double crossing-over, i. e., from a three-locus experiment.
The most accurate data regarding the value for reduplicated and vermilion show that reduplicated is 1.7 units away from vermilion, either to the right or to the left. The distance from white is 29, which means the location for reduplication would be to the left of vermilion, which is at 33. The data for bar indicate a distance of 21, but since bar is itself 24 units from vermilion, this distance of 21 seems to place the location to the right of vermilion. The evidence slightly favors this position to the right of vermilion at 34.7, where reduplicated may be placed provisionally. In any case, the location is so close to that of vermilion that the final decision must come from data involving double crossing-over, i.e., from a three-locus experiment.
LETHAL 1.
DEADLY 1.
In February 1912 Miss E. Rawls found that certain females from a wild stock were giving only about half as many sons as daughters. Tests continuing through five generations showed that the sons that appeared were entirely normal, but that half of the daughters gave again 2 : 1 sex-ratios, while the other half gave normal 1 : 1 sex-ratios. {32}
In February 1912, Miss E. Rawls discovered that certain females from a wild stock were producing only about half as many sons as daughters. Tests conducted over five generations revealed that the sons that were born were completely normal, but half of the daughters produced 2:1 sex ratios, while the other half had normal 1:1 sex ratios. {32}
The explanation of this mode of transmission became clear when it was found that the cause of the death of half of the males was a particular factor that had as definite a locus in the X chromosome as have other sex-linked factors (Morgan, 1912e). Morgan mated females (from the stock sent to him by Miss Rawls) to white-eyed males. Half of the females, as expected, gave 2 : 1 sex-ratios, and daughters from these were again mated to white males. Here once more half of the daughters gave 2 : 1 sex-ratios, but in such cases the sons were nearly all white-eyed and only rarely a red-eyed son appeared, when under ordinary circumstances there should be just as many red sons as white sons. The total output for 11 such females was as follows (Morgan, 1914b): white ♀, 457; red ♀, 433; white ♂, 370; red ♂, 2. It is evident from these data that there must be present in the sex-chromosome a gen that causes the death of every male that receives this chromosome, and that this lethal factor lies very close to the factor for white eyes. The linkage of this lethal (now called lethal 1) to various other sex-linked gens was determined (Morgan 1914b), and is summarized in table 5. On the basis of these data it is found that the gen lethal 1 lies 0.4 unit to the left of white, or at 0.7.
The explanation of this mode of transmission became clear when it was discovered that the cause of death for half of the males was a specific factor that had a clear location on the X chromosome, just like other sex-linked factors (Morgan, 1912e). Morgan crossed females (from the stock sent to him by Miss Rawls) with white-eyed males. As expected, half of the females produced a 2:1 sex ratio, and the daughters from these were again crossed with white males. Once more, half of the daughters produced a 2:1 sex ratio, but in these cases, nearly all of the sons were white-eyed, and only a few red-eyed sons appeared, when normally there should have been just as many red sons as white sons. The total output for 11 such females was as follows (Morgan, 1914b): white ♀, 457; red ♀, 433; white ♂, 370; red ♂, 2. These data clearly indicate that there must be a gene present in the sex chromosome that causes the death of every male that inherits this chromosome, and this lethal factor is located close to the factor for white eyes. The linkage of this lethal factor (now called lethal 1) to various other sex-linked genes was determined (Morgan 1914b) and is summarized in table 5. Based on this data, it was found that the gene lethal 1 is positioned 0.4 unit to the left of white, or at 0.7.
Table 5.—Summary of linkage data upon lethal 1, from Morgan, 1914b, pp. 81-92.
Table 5.—Summary of linkage data on lethal 1, from Morgan, 1914b, pp. 81-92.
| Gens. | Total. | Cross- overs. | Cross-over values. |
| Yellow lethal 1 | 131 | 1 | 0.8 |
| Yellow miniature | 131 | 45 | 34.4 |
| Lethal 1 white | 1,763 | 7 | 0.4 |
| Lethal 1 miniature | 814 | 323 | 39.7 |
| White miniature | 994 | 397 | 39.9 |
LETHAL 1a.
LETHAL 1a.
In the second generation of the flies bred by Miss Rawls, one female gave (March 1912) only 3 sons, although she gave 312 daughters. It was not known for some time (see lethals 3 and 3a) what was the cause of this extreme rarity of sons. It is now apparent, however, that this mother carried lethal 1 in one X and in the other X a new lethal which had arisen by mutation. The new lethal was very close to lethal 1, as shown by the rarity of the surviving sons, which are cross-overs between lethal 1 and the new lethal that we may call lethal 1a. There is another class of cross-overs, namely, those which have lethal 1 and get lethal 1a by crossing-over. These doubly lethal males must also die, but since they are theoretically as numerous as the males (3) free from both lethals, we must double this number (3 × 2) to get the total number of cross-overs. There were 312 daughters, but as the sons are normally about 96 per cent of the number of the females, {33}we may take 300 as the number of the males which died. There must have been, then, about 2 per cent of crossing-over, which makes lethal 1a lie about 2 units from lethal 1. This location of lethal 1a is confirmed by a test that Miss Rawls made of the daughters of the high-ratio female. Out of 98 of these daughters none repeated the high sex-ratio and only 2 gave 1 ♀ : 1 ♂ ratios. The two daughters which gave 1 : 1 ratios are cross-overs. There should be an equal number of cross-overs which contain both lethals. These latter would not be distinguishable from the non-cross-over females, each of which carries one or the other lethal. In calculation, allowance can be made for them by doubling the number of observed cross-overs (2 × 2) and taking 98 - 2 as the number of non-cross-overs. The cross-over fraction {6 + 4}/{300 + 96} gives 2.6 as the distance between the two lethals. Lethal 1a is probably to the right of lethal 1 at 0.7 + 2.6 = 3.3.
In the second generation of flies bred by Miss Rawls, one female (March 1912) produced only 3 males but had 312 daughters. For a while, it was unclear (see lethals 3 and 3a) why there were so few sons. It is now clear that this mother carried lethal 1 on one X chromosome and a new lethal that arose from mutation on the other X. The new lethal is very close to lethal 1, as indicated by the low number of surviving males, which are crossovers between lethal 1 and the new lethal we can call lethal 1a. There’s another type of crossover: those that carry lethal 1 and acquire lethal 1a through crossing over. These doubly lethal males must also die, but since they are theoretically as numerous as the males (3) unaffected by both lethals, we need to double this number (3 × 2) to find the total number of crossovers. There were 312 daughters, but since the number of sons is usually about 96 percent of the number of females, {33} we can consider that 300 males must have died. This indicates there was about 2 percent of crossing-over, placing lethal 1a approximately 2 units away from lethal 1. This position of lethal 1a is supported by a test Miss Rawls conducted on the daughters of the high-ratio female. Among 98 of these daughters, none exhibited the high sex ratio, and only 2 produced a 1 ♀ : 1 ♂ ratio. The two daughters that produced 1:1 ratios are crossovers. There should be an equal number of crossovers that include both lethals. These could not be distinguished from the non-crossover females, each carrying one of the lethals. In calculations, adjustments can be made for them by doubling the observed crossovers (2 × 2) and using 98 - 2 as the count of non-crossovers. The crossover fraction {6 + 4}/{300 + 96} yields 2.6 as the distance between the two lethals. Lethal 1a is likely positioned to the right of lethal 1 at 0.7 + 2.6 = 3.3.
SPOT.
Spot.
(Plate II, figures 14 to 17.)
(__A_TAG_PLACEHOLDER_0__, figures 14-17.)
In April 1912 there was found in the stock of yellow flies a male that differed from yellow in that it had a conspicuous light spot on the upper surface of the abdomen (Morgan, 1914a). In yellow flies this region is dark brown in color. In crosses with wild flies the spot remained with the yellow, and although some 30,000 flies were raised, none of the gray offspring showed the spot, which should have occurred had crossing-over taken place. The most probable interpretation of spot is that it was due to another mutation in the yellow factor, the first mutation being from gray to yellow and the second from yellow to spot.
In April 1912, a male was discovered among the stock of yellow flies that had a noticeable light spot on the upper surface of its abdomen (Morgan, 1914a). In yellow flies, this area is dark brown. When crossed with wild flies, the spot stayed with the yellow, and even though around 30,000 flies were bred, none of the gray offspring had the spot, which should have appeared if crossing-over had occurred. The most likely explanation for the spot is that it was caused by another mutation in the yellow gene, with the first mutation changing gray to yellow and the second changing yellow to the spot.
Spot behaves as an allelomorph to yellow in all crosses where the two are involved and is completely recessive to yellow, i. e., the yellow-spot hybrid is exactly like yellow. A yellow-spot female, back-crossed to a spot male, produces yellows and spots in equal numbers.
Spot acts as an allele to yellow in all crosses where both are present and is fully recessive to yellow, i.e., the yellow-spot hybrid looks exactly like yellow. A yellow-spot female, when bred back to a spot male, produces yellows and spots in equal amounts.
In a cross of spot to black it was found that the double recessive, spot black, flies that appear in F2 have, in addition to the spot on the abdomen, another spot on the scutellum and a light streak on the thorax. These two latter characters ("dot and dash") are very sharply marked and conspicuous when the flies are young, but they are only juvenile characters and disappear as the flies become older. The spot flies never show the "dot and dash" clearly, and it only comes out when black acts as a developer. These characters furnish a good illustration of the fact that mutant gens ordinarily affect many parts of the body, though these secondary effects often pass unnoticed.
In a cross of spot to black, it was found that the double recessive, spot black, flies appearing in F2 not only have a spot on their abdomen but also another spot on their scutellum and a light streak on their thorax. These last two features ("dot and dash") are very distinct and noticeable when the flies are young, but they are only juvenile traits and fade as the flies get older. The spot flies never display the "dot and dash" clearly, and it only appears when black acts as a developer. These traits provide a good example of how mutant genes typically impact multiple body parts, even though these secondary effects often go unnoticed.
In the F2 of the cross of spot by black one yellow black fly appeared, although none are expected, on the assumption that spot and yellow {34}are allelomorphic. Unless due to crossing-over it must have been a mutation from spot back to yellow. Improbable as this may seem to those who look upon mutations as due to losses from the germ-plasm, yet we have records of several other cases where similar mutations "backwards" have taken place, notably in the case of eosin to white, under conditions where the alternative interpretation of crossing-over is excluded.
In the F2 generation from the cross of the spotted and black parent, a yellow-black fly appeared, even though none were expected, based on the assumption that the spotted and yellow are allelomorphic. Unless it was due to crossing-over, it must have been a mutation from spotted back to yellow. As unlikely as this may seem to anyone who thinks mutations result from losses in the germ-plasm, there are several records of similar "backward" mutations occurring, especially in the case of eosin turning to white, under conditions that rule out the alternative explanation of crossing-over. {34}
SABLE.
SABLE.
(Plate I, figure 2.)
(__A_TAG_PLACEHOLDER_0__, fig. 2.)
In an experiment involving black body-color[3] a fly appeared (July 19, 1912) whose body-color differed slightly from ordinary black in that the trident mark on the thorax was sharper and the color itself was brighter and clearer. This fly, a male, was mated to black females and gave some black males and females, but also some gray (wild body-color) males and females, showing not only that he was heterozygous for ordinary recessive black, but at the same time that his dark color must be due to another kind of black. The gray F1 flies when mated together gave a series of gray and dark flies in F2 about as follows: In the females 3 grays to 1 dark; in the males 3 grays to 5 dark in color. The result indicated that the new black color, which we call sable, was due to a sex-linked factor. It was difficult to discover which of the heterogeneous F2 males were the new blacks. Suspected males were bred (singly) to wild females, and the F2 dark males, from those cultures that gave the closest approach to a 2 gray ♀ : 1 gray ♂ : 1 dark ♂, were bred to their sisters in pairs in order to obtain sable females and males. Thus stock homozygous for sable but still containing black as an impurity was obtained. It became necessary to free it from black by successive individual out-crossings to wild flies and extractions.
In an experiment involving black body color, a fly appeared (July 19, 1912) whose body color was slightly different from regular black in that the trident mark on the thorax was sharper, and the color itself was brighter and clearer. This male fly was mated with black females and produced some black males and females, but also some gray (wild body color) males and females, showing that he was heterozygous for the ordinary recessive black, and that his dark color must be due to another type of black. The gray F1 flies, when mated together, produced a series of gray and dark flies in F2 as follows: In the females, there were 3 grays for every 1 dark; in the males, there were 3 grays for every 5 dark. The results indicated that the new black color, which we call sable, was due to a sex-linked factor. It was challenging to determine which of the heterogeneous F2 males were the new blacks. Suspected males were bred (one at a time) to wild females, and the F2 dark males from those pairings that came closest to a ratio of 2 gray ♀ : 1 gray ♂ : 1 dark ♂ were bred to their sisters in pairs to produce sable females and males. This process resulted in a stock that was homozygous for sable but still contained some black as an impurity. It became necessary to eliminate the black through successive individual out-crossings to wild flies and extractions.
This account of how sable was purified shows how difficult it is to separate two recessive factors that give closely similar somatic effects. If a character like sable should be present in any other black stock, or if a character like black should be present in sable, very erratic results would be obtained if such stocks were used in experiments, before such a population had been separated into its component races.
This explanation of how sable was purified illustrates how hard it is to separate two recessive factors that produce very similar physical traits. If a trait like sable showed up in any other black lineage, or if a trait like black appeared in sable, the outcomes would be unpredictable if those lineages were used in experiments, before the population was divided into its distinct races.
Sable males of the purified stock were mated to wild females and gave wild-type (gray) males and females. These inbred gave the results shown in table 6.
Sable males from the pure lineage were mated with wild females, resulting in wild-type (gray) males and females. These inbred produced the outcomes displayed in table 6.
No sable females appeared in F2, as seen in table 6. The reciprocal cross gave the results shown in table 7.
No sable females appeared in F2, as shown in table 6. The reciprocal cross produced the results displayed in table 7.
The F1 males were sable like their mother. The evidence thus shows that sable is a sex-linked recessive character. Our next step was to determine the linkage relations of sable to certain other sex-linked gens, namely, yellow, eosin, cherry, vermilion, miniature, and bar.
The F1 males were sable like their mother. This evidence shows that sable is a sex-linked recessive trait. Our next step was to figure out how sable is linked to other sex-linked genes, specifically yellow, eosin, cherry, vermilion, miniature, and bar.
Table 6.—P1 wild ♀ ♀ × sable ♂. F1 wild-type ♀ ♀ × F1 wild-type ♂ ♂.
Table 6.—P1 wild females × sable males. F1 wild-type females × F1 wild-type males.
| Reference.[4] | Wild-type ♀. | Wild-type ♂. | Sable ♂. |
| 88 C | 218 | 100 | 70 |
| 143 C | 245 | 108 | 72 |
| 146 C | 200 | 115 | 82 |
| Total | 663 | 323 | 224 |
Table 7.—P1 sable ♀ × wild ♂ ♂. F1 wild-type ♀ × F1 sable ♂.
Table 7.—P1 sable female × wild male males. F1 wild-type female × F1 sable male.
| Reference. | Wild-type ♀. | Wild-type ♂. | Sable ♀. | Sable ♂. |
| 4 I | 10 | 10 | 6 | 10 |
LINKAGE OF YELLOW AND SABLE.
LINKING YELLOW AND TAN.
The factor for yellow body-color lies at one end of the known series of sex-linked gens. As already stated, we speak of this end as the left end of the diagram, and yellow as the zero in locating factors.
The factor for yellow body color is at one end of the known series of sex-linked genes. As mentioned earlier, we refer to this end as the left end of the diagram, and yellow as the baseline for locating factors.
When yellow (not-sable) females were mated to (not-yellow) sable males they gave wild-type (gray) daughters and yellow sons. These inbred gave in F2 two classes of females, namely, yellow and gray, and four classes of males, namely, yellow and sable (non-cross-overs), wild type and the double recessive yellow sable (cross-overs). From off-spring (F3) of the F2 yellow sable males by F2 yellow females, pure stock of the double recessive yellow sable was made up and used in the crosses to test linkage.
When yellow (not-sable) females were bred with (not-yellow) sable males, they produced wild-type (gray) daughters and yellow sons. These inbred offspring produced in F2 two types of females, which were yellow and gray, and four types of males, which included yellow and sable (non-cross-overs), wild type, and the double recessive yellow sable (cross-overs). From the offspring (F3) of the F2 yellow sable males and F2 yellow females, a pure stock of the double recessive yellow sable was created and used in crosses to test linkage.
In color the yellow sable is quite similar to yellow black, that is, a rich brown with a very dark brown trident pattern on the thorax. Yellow sable is easier to distinguish from yellow than is yellow black, even when the flies have not yet acquired their adult body-color.
In color, yellow sable closely resembles yellow black, which is a rich brown with a very dark brown trident pattern on the thorax. Yellow sable is easier to tell apart from yellow than yellow black is, even when the flies haven't yet developed their adult body color.
Table 8.—P1 wild ♀ ♀ × yellow sable ♂ ♂. F1 wild-type ♀ ♀ × F1 wild-type ♂ ♂.
Table 8.—P1 wild female × yellow sable male. F1 wild-type female × F1 wild-type male.
| Reference. | Wild- type ♀. | Non-cross-over ♂. | Cross-over ♂. | Total males. | Cross-over value. | ||
| Yellow sable. | Wild- type. | Yellow. | Sable. | ||||
| 44 I | 292 | 110 | 43 | 75 | 36 | 264 | 42 |
| 45 I | 384 | 104 | 58 | 71 | 60 | 293 | 45 |
| Total | 676 | 214 | 101 | 146 | 96 | 557 | 43 |
Some of the F1 females were back-crossed to yellow sable males and gave the data for table 9.
Some of the F1 females were bred back to yellow sable males, which provided the data for table 9.
Table 9.—P1 wild-type ♀ ♀ × yellow sable ♂ ♂. B. C. F1 wild-type ♀ × yellow sable ♂ ♂.
Table 9.—P1 wild-type female × yellow sable male. B. C. F1 wild-type female × yellow sable male.
| Reference. | Non-cross-overs. | Cross-overs. | Total. | Cross-over value. | ||
| Wild-type. | Yellow sable. | Yellow. | Sable. | |||
| 31 I | 108 | 51 | 58 | 56 | 273 | 42 |
| 49 I | 265 | 175 | 161 | 169 | 770 | 43 |
| Total | 373 | 226 | 219 | 225 | 1,043 | 43 |
In these tables the last column (to the right) shows for each culture the amount of crossing-over between yellow and sable. These values are found by dividing the number of cross-overs by the total number of individuals which might show crossing-over, that is, males only or both males and females, as the case may be. Free assortment would give 50 per cent of cross-overs and absolute linkage 0 per cent of cross-overs. Except where the percentage of crossing-over is very small these values are expressed to the nearest unit, since the experimental error might make a closer calculation misleading.
In these tables, the last column (on the right) shows the amount of crossing-over between yellow and sable for each culture. These values are calculated by dividing the number of cross-overs by the total number of individuals that could show crossing-over, which includes either males only or both males and females, depending on the situation. Free assortment would result in 50 percent of cross-overs, while absolute linkage would result in 0 percent of cross-overs. Unless the percentage of crossing-over is very small, these values are rounded to the nearest whole number, as the experimental error might make a more precise calculation misleading.
The combined data of tables 8 and 9 give 686 cross-overs in a total of 1,600 individuals in which crossing-over might occur. The females of table 8 are all of one class (wild type) and are useless for this calculation except as a check upon viability. The cross-over value of 43 per cent shows that crossing-over is very free. We interpret this to mean that sable is far from yellow in the chromosome. Since yellow is at one end of the known series, sable would then occupy a locus somewhere near the opposite end. This can be checked up by finding its linkage relations to the other sex-linked factors. {37}
The combined data from tables 8 and 9 show 686 cross-overs among a total of 1,600 individuals where crossing over could happen. The females in table 8 are all the same type (wild type) and aren't useful for this calculation apart from verifying viability. The cross-over rate of 43 percent indicates that crossing over happens quite freely. We understand this to mean that sable is significantly distant from yellow on the chromosome. Since yellow is positioned at one end of the known series, sable would then be located near the opposite end. This can be validated by examining its linkage relationships with the other sex-linked factors. {37}
LINKAGE OF CHERRY AND SABLE.
Linking Cherry and Sable.
The origin of cherry eye-color (Plate II, fig. 9) has been given by Safir (Biol. Bull., 1913). From considerations which will be discussed later in this paper we regard cherry as allelomorphic to white in a quadruple allelomorph system composed of white, eosin, cherry, and their normal red allelomorph. Cherry will then occupy the same locus as white, which is one unit to the right of yellow, and will show the same linkage relations to other factors as does white. A slightly lower cross-over value should be given by cherry and sable than was given by yellow and sable.
The origin of cherry eye color (Plate II, fig. 9) was discussed by Safir (Biol. Bull., 1913). Based on points that will be covered later in this paper, we consider cherry to be allelomorphic to white in a four-type allelomorph system that includes white, eosin, cherry, and their normal red allelomorph. Therefore, cherry will occupy the same locus as white, which is positioned one unit to the right of yellow, and will exhibit the same linkage relationships to other factors as white does. Cherry and sable should have a slightly lower cross-over value than what was observed with yellow and sable.
When cherry (gray) females were crossed to (red) sable males the daughters were wild type and the sons cherry. Inbred these gave the results shown in table 10.
When cherry (gray) females were mated with (red) sable males, the daughters were wild type and the sons were cherry. Inbreeding these produced the results shown in table 10.
Table 10.—P1 cherry ♀♀ × sable ♂♂. F1 wild-type ♀ × F1 cherry ♂ ♂.
Table 10.—P1 cherry females × sable males. F1 wild-type females × F1 cherry males.
| Reference. | Wild- type ♀. | Cherry ♀. | Non-cross- over ♂. | Cross-over ♂. | Total males. | Cross- over value. | ||
| Cherry. | Sable. | Cherry sable. | Wild- type. | |||||
| 24 I | 94 | 105 | 51 | 42 | 20 | 43 | 156 | 40 |
| 55 I | 101 | 131 | 63 | 52 | 38 | 48 | 201 | 43 |
| 55′ I | 96 | 94 | 52 | 31 | 29 | 30 | 142 | 42 |
| Total | 291 | 330 | 166 | 125 | 87 | 121 | 499 | 42 |
The percentage of crossing-over between cherry and sable is 42. Since cherry is one point from yellow, this result agrees extremely well with the value 43 for yellow and sable. Since yellow and eosin lie at the left end of the first chromosome, the high values, namely, 43 and 42, agree in making it very probable that sable lies near the other end (i. e., to the right). Sable will lie farther to the right than vermilion, for vermilion has been shown elsewhere to give 33 per cent of crossing-over with eosin. The location of sable to the right of vermilion has in fact been substantiated by all later work.
The percentage of crossing-over between cherry and sable is 42. Since cherry is one point away from yellow, this result closely matches the value of 43 for yellow and sable. Given that yellow and eosin are located at the left end of the first chromosome, the high values of 43 and 42 suggest that sable is likely positioned near the other end (i.e., to the right). Sable will be further to the right than vermilion, as it has been shown in other studies that vermilion produces a 33 percent crossing-over rate with eosin. The positioning of sable to the right of vermilion has indeed been confirmed by all subsequent research.
LINKAGE OF EOSIN, VERMILION, AND SABLE.
LINKAGE OF EOSIN, VERMILION, AND SABLE.
Three loci are involved in the next experiment. Since eosin is an allelomorph of cherry, it should be expected to give with sable the same cross-over value as did cherry. When eosin (red) sable females were crossed to (red) vermilion (gray) males, the daughters were wild type and the males were eosin sable. Inbred these gave the classes shown in table 11. {38}
Three locations are involved in the next experiment. Since eosin is an allelomorph of cherry, it is expected to produce the same crossover value with sable as cherry did. When eosin (red) sable females were crossed with (red) vermilion (gray) males, the daughters were wild type and the males were eosin sable. Inbreeding these resulted in the classes shown in table 11. {38}
Table 11.—P1 eosin sable ♀ × vermilion ♂♂. F1 wild-type ♀♀ × F1 eosin sable ♂♂.
Table 11.—P1 eosin sable female × vermilion males. F1 wild-type females × F1 eosin sable males.
| Reference. | F2 females. | F2 males. | ||||||||||

| 
| 
| 
| 
|  | |||||||
| Eosin sable. | Wild- type. | Eosin. | Sable. | Eosin sable. | Ver- milion. | Eosin ver- milion. | Sable. | Eosin. | Ver- milion sable. | Eosin ver- milion sable. | Wild- type. | |
| 26 I | 132 | 171 | 113 | 109 | 127 | 163 | 75 | 76 | 37 | 14 | 2 | 5 |
| 26′I | 96 | 146 | 86 | 78 | 74 | 128 | 76 | 59 | 18 | 21 | 4 | 3 |
| Total. | 228 | 317 | 199 | 187 | 201 | 291 | 151 | 135 | 55 | 35 | 6 | 8 |
If we consider the male classes of table 11, we find that the smallest classes are eosin vermilion sable and wild type, which are the expected double cross-over classes if sable lies to the right of vermilion, as indicated by the crosses with eosin and with yellow. The classes which represent single crossing-over between eosin and vermilion are eosin vermilion, and sable, and those which represent single crossing-over between vermilion and sable are eosin and vermilion sable. These relations are seen in diagram II.
If we look at the male groups in table 11, we see that the smallest groups are eosin vermilion sable and wild type, which are the expected double crossover groups if sable is to the right of vermilion, as shown by the crosses with eosin and yellow. The groups that show single crossing-over between eosin and vermilion are eosin vermilion and sable, while the groups that show single crossing-over between vermilion and sable are eosin and vermilion sable. You can see these relationships in diagram II.

Diagram II.—The upper line represents an X chromosome, the lower line its mate. The cross connecting lines indicate crossing-over between pairs of factors.
Diagram II.—The upper line shows an X chromosome, while the lower line shows its pair. The connecting lines indicate crossing-over between pairs of factors.
| Non-cross-overs |  |  | Eosin sable. |
| Vermilion. | |||
| Single cross-overs |  | | Eosin vermilion. |
| Sable. | |||
 | | Eosin. | |
| Vermilion sable. | |||
| Double cross-overs |  | | Eosin vermilion sable. |
| Wild-type. |
If we consider the female classes of table 11, we get information as to the cross-over value of eosin and sable, namely, 42 units. The male classes will be considered in connection with the cross that follows.
If we look at the female categories in table 11, we see that the cross-over value of eosin and sable is 42 units. We'll examine the male categories in relation to the upcoming cross.
The next experiment involves the same three gens which now enter in different relations. A double recessive, eosin vermilion (gray) female {39}was mated to (red red) sable males and gave 202 wild-type[5] females and 184 eosin vermilion males. Two F1 pairs gave the results shown in table 12 (the four classes of females not being separated).
The next experiment involves the same three genes, which now interact in different ways. A double recessive eosin vermilion (gray) female {39}was mated with (red red) sable males and produced 202 wild-type[5] females and 184 eosin vermilion males. Two F1 pairs produced the results shown in table 12 (the four classes of females were not separated).
Table 12.—P1 eosin vermilion F1 wild-type ♀ × F1 eosin vermilion ♂ ♂.
Table 12.—P1 eosin vermilion F1 wild-type female × F1 eosin vermilion male male.
| Reference. | F2 females. | F2 males. | |||||||

| 
| 
|  | ||||||
| Eosin Ver- milion ♂. | Sable ♂. | Eosin sable ♂. | Ver- milion. ♂. | Eosin Ver- milion sable ♂. | Wild- type ♂. | Eosin ♂. | Ver- milion sable ♂. | ||
| 59 C | 133 | 40 | 33 | 7 | 16 | 5 | 5 | 2 | 1 |
| 61 C | 101 | 34 | 26 | 8 | 11 | 3 | 7 | 1 | 0 |
| Total | 234 | 74 | 59 | 15 | 27 | 8 | 12 | 3 | 1 |
If we combine the data for males given in table 12 with those of table 11, we get the following cross-over values. Eosin vermilion, 32; vermilion sable, 12; eosin sable, 41.
If we combine the data for males in table 12 with the data from table 11, we get the following crossover values: eosin vermilion, 32; vermilion sable, 12; eosin sable, 41.
LINKAGE OF MINIATURE AND SABLE.
LINKING MINIATURE AND SABLE.
The miniature wing has been described (Morgan, Science, 1911) and the wing figured (Morgan, Jour. Exp. Zool., 1911). The gen for miniature lies about 3 units to the right of vermilion, so that it is still closer to sable than is vermilion. The double recessive, miniature sable, was made up, and males of this stock were bred to wild females (long gray). The wild-type daughters were back-crossed to double recessive males and gave the results (mass cultures) shown in table 13.
The tiny wing has been discussed (Morgan, Science, 1911) and the wing was illustrated (Morgan, Jour. Exp. Zool., 1911). The gene for tiny wings is located about 3 units to the right of vermilion, making it closer to sable than vermilion is. The double recessive, tiny sable, was created, and males from this line were bred with wild females (long gray). The wild-type daughters were back-crossed with double recessive males, resulting in the outcomes (mass cultures) shown in table 13.
Table 13.—P1 wild ♀ ♀ × miniature sable ♂ ♂. B. C. F1 wild-type ♀ ♀ × miniature sable ♂ ♂.
Table 13.—P1 wild females × miniature sable males. B. C. F1 wild-type females × miniature sable males.
| Reference. | Non-cross-overs. | Cross-overs. | Total. | Cross- over value. | ||
| Miniature sable. | Wild-type. | Miniature. | Sable. | |||
| 38 I | 245 | 283 | 15 | 17 | 560 | 6 |
| 43 I | 191 | 236 | 13 | 18 | 458 | 7 |
| 46 I | 232 | 274 | 24 | 21 | 551 | 8 |
| Total | 668 | 793 | 52 | 56 | 1,569 | 7 |
Since the results for the male and the female classes are expected to be the same, the sexes were not separated. The combined data give 7 per cent of crossing-over between miniature and sable.
Since the results for the male and female groups are expected to be the same, the sexes were not separated. The combined data shows a 7 percent occurrence of crossing-over between miniature and sable.
LINKAGE OF VERMILION, SABLE, AND BAR.
LINKAGE OF VERMILION, SABLE, AND BAR.
Bar eye has been described by Mrs. S. C. Tice (1914). It is a dominant sex-linked character, whose locus, lying beyond vermilion and sable, is near the right end of the chromosome series, that is, at the end opposite yellow.
Bar eye has been described by Mrs. S. C. Tice (1914). It is a dominant sex-linked trait, with its location lying beyond vermilion and sable, near the end of the chromosome series, specifically at the end opposite yellow.
In the first cross of a balanced series of experiments for the gens vermilion, sable, and bar, vermilion (gray not-bar) entered from one side (♀) and (red) sable bar from the other (♂). The daughters were bar and the sons vermilion. The daughters were back-crossed singly to the triple recessive males vermilion sable (not-bar), and gave the data included in table 14.
In the first round of a balanced series of experiments for the gens vermilion, sable, and bar, vermilion (gray not-bar) came in from one side (♀) and (red) sable bar from the other side (♂). The daughters were bar and the sons were vermilion. The daughters were back-crossed individually to the triple recessive males vermilion sable (not-bar), resulting in the data included in table 14.
In the second cross, vermilion sable (not-bar) went in from one side (♀) and (red, gray) bar from the other. The daughters were bar and the sons were vermilion sable. Since these sons have the three recessive factors, inbreeding of F1 is equivalent to a triple back-cross. The results are given by pairs in table 15. {41}
In the second cross, vermilion sable (not-bar) came in from one side (♀) and (red, gray) bar from the other. The daughters were bar and the sons were vermilion sable. Since these sons carry three recessive factors, inbreeding of F1 is like doing a triple back-cross. The results are provided in pairs in table 15. {41}
Table 14.—P1 vermilion ♀ ♀ × sable bar ♂ ♂. B. C. F1 bar ♀ × vermilion sable ♂ ♂.
Table 14.—P1 vermilion female × sable bar male. B. C. F1 bar female × vermilion sable male.
| Reference. | 
| 
| 
| 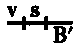
| Total. | Cross-over values. | ||||||
| Ver- milion. | Sable bar. | Ver- milion sable bar. | Wild- type. | Ver- milion bar. | Sable. | Ver- milion sable. | Bar. | Ver- milion sable. | Sable bar. | Ver- milion bar. | ||
| 147 I | 81 | 66 | 12 | 15 | 15 | 18 | 207 | 13 | 16 | 29 | ||
| 148 I | 103 | 108 | 4 | 19 | 11 | 11 | 256 | 9 | 9 | 18 | ||
| 149 I | 97 | 88 | 10 | 8 | 17 | 17 | 1 | 1 | 239 | 8 | 15 | 22 |
| 150 I | 95 | 75 | 10 | 11 | 21 | 22 | 1 | 1 | 236 | 10 | 19 | 27 |
| 151 I | 116 | 96 | 11 | 15 | 23 | 26 | 2 | 289 | 10 | 18 | 26 | |
| 89 | 89 | 94 | 10 | 19 | 15 | 11 | 1 | 239 | 13 | 11 | 23 | |
| 90 | 49 | 50 | 4 | 8 | 15 | 14 | 140 | 9 | 21 | 29 | ||
| 91 | 104 | 88 | 13 | 15 | 12 | 12 | 244 | 11 | 10 | 21 | ||
| Total. | 734 | 665 | 74 | 110 | 129 | 131 | 3 | 4 | 1,850 | 10 | 14 | 24 |
Table 15.—P1 vermilion sable ♀ ♀ × bar ♂ ♂. B. C. F1 bar ♀ × vermilion sable ♂ ♂.
Table 15.—P1 vermilion sable female × bar male. B. C. F1 bar female × vermilion sable male.
| Reference. | 
| 
| 
| 
| Total. | Cross-over values. | ||||||
| Ver- milion sable. | Bar | Ver- milion bar. | Sable. | Ver- milion sable bar. | Wild- type. | Ver- milion. | Sable bar. | Ver- milion sable. | Sable bar. | Ver- milion bar. | ||
| 105 I | 41 | 75 | 10 | 4 | 5 | 11 | 146 | 10 | 11 | 21 | ||
| 106 I | 59 | 122 | 16 | 13 | 11 | 17 | 238 | 12 | 12 | 24 | ||
| 107 I | 92 | 98 | 8 | 12 | 16 | 10 | 236 | 9 | 11 | 20 | ||
| 116 I | 111 | 149 | 19 | 16 | 20 | 19 | 1 | 335 | 11 | 12 | 22 | |
| 117 I | 92 | 117 | 16 | 14 | 15 | 18 | 272 | 11 | 12 | 23 | ||
| 126 I | 96 | 160 | 13 | 13 | 17 | 35 | 334 | 8 | 15 | 23 | ||
| 127 I | 117 | 124 | 13 | 25 | 24 | 30 | 1 | 334 | 12 | 16 | 28 | |
| Total | 608 | 845 | 95 | 97 | 108 | 140 | 1 | 1 | 1,895 | 10 | 13 | 23 |
In the third cross, vermilion (gray) bar entered from one side (♀) and (red) sable (not-bar) from the other (♂). The daughters are bar and the sons vermilion bar. The daughters were back-crossed singly to vermilion sable males and gave the data in table 16.
In the third cross, the gray bar came in from one side (♀) and the red sable (not-bar) came from the other (♂). The daughters are bar and the sons have vermilion bar. The daughters were back-crossed individually to vermilion sable males, and the results are shown in table 16.
Table 16.—P1 vermilion bar ♀ ♀ × sable ♂ ♂. B. C. F1 bar ♀ × vermilion sable ♂ ♂.
Table 16.—P1 vermilion bar female × sable male. B. C. F1 bar female × vermilion sable male.
| Reference. | 
| 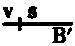
| 
| 
| Total. | Cross-over values. | ||||||
| Ver- milion bar. | Sable. | Ver- milion sable. | Bar. | Ver- milion. | Sable bar. | Ver- milion sable bar. | Wild- type. | Ver- milion sable. | Sable bar. | Ver- milion bar. | ||
| 129 I | 132 | 147 | 15 | 15 | 19 | 21 | 1 | 1 | 351 | 9 | 12 | 20 |
| 130 I | 194 | 168 | 21 | 17 | 28 | 25 | .. | 1 | 454 | 9 | 12 | 20 |
| 131 I | 121 | 89 | 10 | 20 | 26 | 11 | 1 | 1 | 279 | 12 | 14 | 24 |
| 137 I | 139 | 113 | 19 | 12 | 33 | 14 | .. | 1 | 331 | 10 | 15 | 24 |
| 138 I | 131 | 128 | 11 | 11 | 28 | 24 | 1 | .. | 334 | 7 | 16 | 22 |
| 139 I | 83 | 79 | 4 | 12 | 17 | 12 | .. | .. | 207 | 8 | 14 | 22 |
| Total. | 800 | 724 | 80 | 87 | 151 | 107 | 3 | 4 | 1,956 | 9 | 14 | 22 |
In the fourth cross, vermilion sable bar entered from one side, and (red gray not-bar) wild type from the other. The daughters were bar and the sons vermilion sable bar. The daughters were back-crossed singly to vermilion sable males, with the results shown in table 17.
In the fourth cross, red-black bar came in from one side, and (red gray wild type) came in from the other. The daughters were bar and the sons were red-black bar. The daughters were back-crossed individually to red-black males, with the results shown in table 17.
Table 17.—P1 vermilion sable bar ♀ ♀ × wild ♂ ♂. B. C. F1 bar ♀ × vermilion sable ♂ ♂.
Table 17.—P1 vermilion sable bar female × wild male. B. C. F1 bar female × vermilion sable male.
| Reference. | 
| 
| 
| 
| Total. | Cross-over values. | ||||||
| Ver- milion sable bar. | Wild- type | Ver- milion. | Sable bar. | Ver- milion sable. | Bar. | Ver- milion bar. | Sable. | Ver- milion sable. | Sable bar. | Ver- milion bar. | ||
| 132 I | 95 | 108 | 10 | 13 | 24 | 22 | .. | .. | 272 | 9 | 17 | 25 |
| 133 I | 112 | 150 | 18 | 16 | 26 | 16 | 1 | 2 | 341 | 11 | 13 | 22 |
| 134 I | 84 | 95 | 14 | 7 | 15 | 16 | .. | 1 | 232 | 10 | 14 | 22 |
| 135 I | 100 | 86 | 16 | 17 | 19 | 22 | .. | 1 | 261 | 13 | 16 | 28 |
| 152 I | 73 | 88 | 12 | 8 | 14 | 18 | .. | .. | 213 | 9 | 15 | 24 |
| 153 I | 114 | 138 | 12 | 12 | 17 | 17 | .. | .. | 310 | 8 | 11 | 19 |
| 154 I | 63 | 90 | 10 | 8 | 8 | 15 | .. | .. | 194 | 9 | 12 | 21 |
| Total. | 641 | 755 | 92 | 81 | 123 | 126 | 1 | 4 | 1,823 | 10 | 14 | 23 |
In tables 14 to 17 the calculations for the three cross-over values for vermilion, sable, and bar are given for the separate cultures and for the totals. The latter are here repeated.
In tables 14 to 17, the calculations for the three cross-over values for vermilion, sable, and bar are provided for the individual cultures and for the totals. The totals are repeated here.
| From— | Vermilion sable. | Sable bar. | Vermilion bar. |
| Table 14 | 10 | 14 | 24 |
| 15 | 10 | 13 | 23 |
| 16 | 9 | 14 | 22 |
| 17 | 10 | 14 | 23 |
The results of the different experiments are remarkably uniform. There can be no doubt that the cross-over value is independent of the way in which the experiment is made, whether any two recessives enter from the same or from opposite sides.
The results of the various experiments are surprisingly consistent. There's no doubt that the cross-over value doesn't depend on how the experiment is conducted, whether two recessives come from the same side or from opposite sides.
Table 18.—Linkage of vermilion, sable, and bar with balanced viability.
Table 18.—Connecting vermilion, sable, and bar with stable viability.

| 
| 
| 
| Total. | |
| Wild-type | 755 | 110 | 140 | 4 | |
| Vermilion | 734 | 92 | 151 | 1 | |
| Sable | 724 | 97 | 131 | 4 | |
| Bar | 845 | 87 | 126 | 4 | |
| Vermilion sable | 608 | 80 | 123 | 3 | |
| Vermilion bar | 800 | 95 | 129 | 1 | |
| Sable bar | 665 | 81 | 107 | 1 | |
| Vermilion sable bar | 641 | 74 | 108 | 3 | |
| Total | 5,772 | 716 | 1,015 | 21 | 7,524 |
| Percentage | 76.7 | 9.53 | 13.49 | 0.28 |
In table 18 the data from each of the four separate experiments have been combined in the manner explained, so that viability is canceled to the greatest extent. The amount of each kind of cross-over appears at the bottom of the table. The total amount of crossing-over between vermilion and sable is the sum of the single (9.53) and of the double (0.28) cross-overs, which value is 9.8. Likewise the cross-over value for sable bar is 13.49 + 0.28 (= 14), and for vermilion bar is 9.53 + 13.49 (= 23). By means of these cross-over values we may calculate the coincidence involved, which is in this case
In Table 18, the data from each of the four separate experiments has been combined as explained, so that viability is minimized as much as possible. The amount of each type of crossover is shown at the bottom of the table. The total amount of crossing-over between vermilion and sable is the sum of the single (9.53) and double (0.28) cross-overs, which totals 9.8. Similarly, the crossover value for the sable bar is 13.49 + 0.28 (= 14), and for the vermilion bar is 9.53 + 13.49 (= 23). Using these crossover values, we can calculate the coincidence involved, which in this case
| 0.0028 × 100 | = 20.8 |
| 0.0953 + 0.0028 × 0.1349 + 0.0028 |
This value shows that there actually occurs only about 21 per cent of the double cross-overs which from the values of the single cross-overs are expected to occur in this section of the chromosome. This is the result which is to be anticipated upon the chromosome view, for if crossing-over is connected with loops of the chromosomes, and if these loops have an average length, then if the chromosomes cross over at one {44}point it is unlikely they will cross over again at another point nearer than the average length of the loop.
This value indicates that only about 21 percent of the double cross-overs happen, which is much lower than what we would expect based on the single cross-overs in this part of the chromosome. This matches what we would expect from the chromosome perspective because if crossing-over is related to the loops in chromosomes, and these loops have an average length, then when chromosomes cross over at one point, it's unlikely they will cross over again at another point that is closer than the average length of the loop. {44}
The calculation of the locus for sable gives 43.0.
The calculation of the locus for sable gives 43.0.
DOT.
DOT.
In the F2, from a cross of a double recessive (white vermilion) female by a triple recessive (eosin vermilion pink) male, there appeared, July 21, 1912, three white-eyed females which had two small, symmetrically placed, black, granular masses upon the thorax. These "dots" appeared to be dried exudations from pores. It did not seem possible that such an effect could be inherited, but as this condition had never been observed before, it seemed worth while to mate the three females to their brothers. In the next generation about 1 per cent of the males were dotted. From these females and males a stock was made up which in subsequent generations showed from 10 to 50 per cent of dot. Selection seemed to have no effect upon the percentage of dot. Although the stock never showed more than 50 per cent of dot, yet it was found that the normal individuals from the stock threw about the same per cent as did those that were dotted, so that the stock was probably genetically pure. The number of males which showed the character was always much smaller than the number of dotted females; in the hatches which produced nearly 50 per cent of dot, nearly all the females but very few of the males were dotted. Quite often the character showed on only one side of the thorax.
In the F2, from a cross of a double recessive (white vermilion) female with a triple recessive (eosin vermilion pink) male, three white-eyed females emerged on July 21, 1912. These females had two small, symmetrically placed black, granular masses on their thorax. These "dots" appeared to be dried secretions from pores. It didn't seem likely that such a trait could be inherited, but since this condition had never been seen before, it was worth mating the three females with their brothers. In the following generation, about 1 percent of the males had dots. A breeding line was established from these females and males, which in later generations showed between 10 and 50 percent dotted individuals. Selection didn't appear to influence the percentage of dotted individuals. Although the breeding line never exceeded 50 percent dotted, it was found that the normal individuals from this line produced about the same percentage as those with dots, suggesting the line was likely genetically pure. The number of males exhibiting the trait was always much lower than the number of dotted females; in the hatches that produced nearly 50 percent dotted, almost all the females but very few of the males were dotted. Often, the trait appeared on only one side of the thorax.
Since this character arose in an experiment involving several eye-colors an effort was made by crossing to wild and extracting to transfer the dot to flies normal in all other respects. This effort succeeded only partly, for a stock was obtained which differed from the wild type only in that it bore dot (about 30 per cent) and in that the eyes were vermilion. Several attempts to get the dot separated from vermilion failed. Since this was only part of the preliminary routine work necessary to get a mutant stock in shape for exact experimentation, no extensive records were kept.
Since this character came from an experiment involving different eye colors, an effort was made by crossing with wild types and extracting to transfer the dot to flies that were normal in every other way. This effort was only partially successful, resulting in a stock that differed from the wild type only by having the dot (about 30 percent) and having vermilion eyes. Multiple attempts to separate the dot from the vermilion color failed. Since this was only a part of the initial routine work needed to prepare a mutant stock for precise experimentation, no detailed records were kept.
LINKAGE OF VERMILION AND DOT.
LINKAGE OF VERMILION AND DOT.
When a dot male with vermilion eyes was bred to a wild female the offspring were wild-type males and females. These inbred gave the data shown in table 19.
When a dot male with red eyes was bred with a wild female, the offspring were wild-type males and females. These inbred produced the data shown in table 19.
Table 19.—P1 vermilion dot ♂ × wild ♀ ♀. F1 wild-type ♀ ♀ × F1 wild-type ♂ ♂.
Table 19.—P1 vermilion dot male × wild female females. F1 wild-type female females × F1 wild-type male males.
| Reference. | F2 females. | Wild-type ♂. | Vermilion ♂. | Vermilion dot ♂. | Dot ♂. |
| 7 | 345 | 151 | 130 | 0 | 0 |
| 8 | 524 | 245 | 220 | 3 | 0 |
| Total. | 869 | 396 | 350 | 3 | 0 |
Only three dot individuals appeared in F2, but since these were males the result indicates that the dot character is due to a sex-linked gen. These three males had also vermilion eyes, indicating linkage of dot and vermilion. The males show no deficiency in numbers, therefore the non-appearance of the dot can not be due to its being semi-lethal. It appears, therefore, that the expression of the character must depend on the presence of an intensifying factor in one of the autosomes, or more probably, like club, it appears only in a small percentage of flies that are genetically pure for the character.
Only three dot individuals showed up in F2, but since these were males, the results suggest that the dot trait is linked to a sex chromosome. These three males also had vermilion eyes, indicating a connection between the dot and vermilion traits. The males don't seem to have a shortage in numbers, so the absence of the dot can't be because it's semi-lethal. It seems, then, that the expression of the trait must rely on the presence of an intensifying factor in one of the autosomes. More likely, similar to club, it shows up only in a small percentage of flies that are genetically pure for the trait.
The reciprocal cross (dot female with vermilion eyes by wild male) was made (table 20). The daughters were wild type and the sons vermilion. Not one of the 272 sons showed dot. If the gen is sex-linked the non-appearance of dot in the F1 males can be explained on the ground that males that are genetically dot show dot very rarely, or that its appearance is dependent upon the intensification by an autosomal factor of the effect produced by the sex-linked factor for dot.
The reciprocal cross (dot female with vermilion eyes by wild male) was made (table 20). The daughters were wild type and the sons were vermilion. Not one of the 272 sons exhibited dot. If the gene is sex-linked, the absence of dot in the F1 males can be explained by the fact that males who are genetically dot show dot very rarely, or that its appearance relies on an autosomal factor intensifying the effect produced by the sex-linked factor for dot.
Table 20.—P1 vermilion dot ♀ × wild ♂.
Table 20.—P1 vermilion dot female × wild male.
| First generation. | Second generation. | |||||||||||
| Reference. | Wild- type ♀. | Ver- milion ♂. | Reference. | Wild- type ♂. | Wild- type ♀. | Ver- milion ♂. | Ver- milion ♀. | Ver- milion dot ♂. | Ver- milion dot ♀. | Dot ♂. | Dot ♀. | |
| 137 C. | 44 | 45 | 19 | 211 | 198 | 228 | 206 | 20 | 3 | 0 | 0 | |
| 138 C. | 77 | 62 | 22 | 266 | 220 | 227 | 227 | 16 | 0 | 0 | 0 | |
| 124 | 124 | 28 | 143 | 149 | 125 | 124 | 14 | 1 | 0 | 0 | ||
| 57 | 41 | Total. | 620 | 567 | 570 | 557 | 50 | 4 | 0 | 0 | ||
| Total. | 291 | 272 | ||||||||||
The F2 generation is given in table 20. The dot reappeared in F2 both in females and in males, but instead of appearing in 50 per cent of both sexes, as expected if it is simply sex-linked, it appeared in 4.0 per cent in the females and in only 0.4 per cent in the males. The failure of the character to be fully realized is again apparent, but here, where it is possible for it to be realized equally in males and females, we find that there are 50 females with dot to only 4 dot males. This would indicate that the character is partially "sex-limited" (Morgan, 1914d) in its realization. The dot appeared only in flies with vermilion eyes, indicating extremely strong linkage between vermilion and dot.
The F2 generation is shown in table 20. The dot reappeared in F2 in both females and males, but instead of showing up in 50 percent of both sexes, as expected if it were simply sex-linked, it appeared in 4.0 percent of females and only in 0.4 percent of males. The incomplete expression of the trait is once again evident, but here, where it could be expressed equally in both males and females, we see that there are 50 females with the dot compared to just 4 males with the dot. This suggests that the trait is partially "sex-limited" (Morgan, 1914d) in its expression. The dot only appeared in flies with vermilion eyes, indicating a very strong linkage between vermilion and dot.
The evidence from the history of the stock, together with these experiments, shows that the character resembles club (wing) in that it is not expressed somatically in all the flies which are homozygous for it. In the case of club we were fortunate enough to find a constant feature {46}which we could use as an index, but, so far as we have been able to see, there is no such constant accessory character in the case of the dot. Unlike club, dot is markedly sex-limited in its effect; that is, there is a difference of expression of the gen in the male and female. This difference recalls the sexual dimorphism of the eosin eye.
The evidence from the history of the stock, along with these experiments, indicates that the trait resembles club (wing) in that it is not physically shown in all flies that are homozygous for it. In the case of club, we were lucky to find a consistent feature {46}that we could use as a reference, but, as far as we can tell, there is no such constant accessory trait in the case of the dot. Unlike club, dot is significantly influenced by sex; that is, the expression of the gene differs between males and females. This difference is reminiscent of the sexual dimorphism seen in the eosin eye.
BOW.
Bend the knee.
In an F2 generation from rudimentary males by wild females there appeared, August 15, 1912, a single male whose wings instead of being flat were turned down over the abdomen (fig. c). The curvature was uniform throughout the length of the wing. A previous mutation, arc, of this same type had been found to be a recessive character in the second group. The new mutation, bow, is less extreme than arc and is more variable in the amount of curvature. When the bow male was mated to wild females the offspring had straight wings.
In an F2 generation from basic males and wild females, a single male was observed on August 15, 1912, whose wings, instead of being flat, curved down over the abdomen (fig. c). The curvature was consistent along the entire length of the wing. A previous mutation, arc, of this same type was identified as a recessive trait in the second group. The new mutation, bow, is less pronounced than arc and shows more variation in the degree of curvature. When the bow male was paired with wild females, the offspring had straight wings.
 Fig. C.—Bow wing.
Fig. C.—Bow wing.
Table 21.—P1 bow ♂♂ × wild ♀♀.
Table 21.—P1 male bow × wild female.
| First generation. | Second generation. | ||||||
| Reference. | Wild-type ♀♀. | Wild-type | Reference. | Wild-type ♀♀. | Wild-type ♂♂. | Bow ♂♂. | |
| 169 C. | 17 | 17 | 18 I. | 193 | 145 | 67 | |
| 21 I | 182 | 100 | 49 | ||||
| Total. | 375 | 245 | 116 | ||||
The F2 ratio in table 21 is evidently the 2:1:1 ratio typical of sex-linkage, but with the bow males running behind expectation. This deficiency is due in part to viability but more to a failure to recognize all the bow-winged individuals, so that some of them were classified among the not-bow or straight wings. In favor of the view that the classification was not strict is the fact that the sum of the two male classes about equals the number of the females.
The F2 ratio in table 21 clearly shows a 2:1:1 ratio, which is typical of sex-linkage, but the bow males are falling short of what was expected. This shortfall is partly due to viability issues and mainly because not all bow-winged individuals were identified, leading some to be categorized as not-bow or straight wings. Supporting the idea that the classification wasn't strict is the fact that the total of the two male categories is roughly equal to the number of females.
BOW BY ARC.
Bend by arc.
When this mutant first appeared its similarity to arc led us to suspect that it might be arc itself or an allelomorph of arc. It was bred, therefore, to arc. The bow male by arc females gave straight (normal) winged males and females. The appearance of straight wings shows that bow is not arc nor allelomorphic to arc. When made later, the reciprocal cross of bow female by arc male gave in F1 straight-winged females but bow males. This result is in accordance with the interpretation that bow is a sex-linked recessive. Further details of these last two experiments may now be given. The F1 (wild-type) flies from bow male by arc female were inbred. The data are given in table 22.
When this mutant first showed up, its resemblance to arc led us to think it might actually be arc or a variant of it. So, we bred it with arc. The offspring from a bow male and arc females produced normal-winged males and females. The presence of normal wings indicates that bow is neither arc nor a variant of it. Later, when we conducted the reciprocal cross of a bow female with an arc male, the F1 offspring were straight-winged females but bow males. This result supports the idea that bow is a sex-linked recessive trait. We can now provide more details about these last two experiments. The F1 (wild-type) flies from the bow male and arc female were inbred. The data can be found in table 22.
Table 22.—P1 bow ♂ × arc ♀.
Table 22.—P1 bow male × arc female.
| First generation. | Second generation. | |||||
| Reference. | Wild-type ♀ ♀. | Wild-type ♂ ♂. | Reference. | Straight. | Not- straight. | |
| 71 C. | 48 | 43 | 71 C. | 179 | 133 | |
| 75 C. | 28 | 27 | ||||
| Total. | 76 | 70 | ||||
Bow and arc are so much alike that they give a single rather variable phenotypic class in F2. Therefore the F2 generation is made up of only two separable classes—flies with straight wings and flies with not-straight wings. The ratio of the two should be theoretically 9:7, which is approximately realized in 179:133.
Bow and arc are so similar that they form one somewhat variable phenotypic category in F2. As a result, the F2 generation consists of just two distinct classes—flies with straight wings and flies with non-straight wings. The expected ratio of these two categories should be 9:7, which is roughly reflected in a ratio of 179:133.
If the distribution of the characters according to sex is ignored, the case is similar to the case of the two white races of sweet peas, which bred together gave wild-type or purple peas in F1 and in F2 gave 9 colored to 7 white. If sex is taken into account, the theoretical expectation for the F2 females is 6 straight to 2 arc, and for the F2 males 3 straight to 1 arc to 3 bow to 1 bow-arc.
If we ignore the distribution of characters by sex, it's similar to the case of the two white varieties of sweet peas, which, when crossed, produced wild-type or purple peas in F1 and resulted in a ratio of 9 colored to 7 white in F2. When we consider sex, the expected ratio for the F2 females is 6 straight to 2 arc, while for the F2 males, it’s 3 straight to 1 arc to 3 bow to 1 bow-arc.
Table 23.—P1 bow ♀ × arc ♂.
Table 23.—P1 female bow × male arc.
| First generation. | Second generation. | |||||
| Reference. | Wild-type ♀ ♀. | Bow ♂ ♂. | Reference. | Straight. | Not- straight. | |
| 72 C. | 22 | 19 | 3 I. | 56 | 69 | |
| 73 C. | 12 | 10 | 3.1 I. | 46 | 62 | |
| 5 I. | 22 | 21 | 5 I. | 56 | 68 | |
| 74 C. | 56 | 52 | 5.1 I. | 90 | 108 | |
| Total. | 112 | 102 | Total. | 248 | 307 | |
In this case the F2 expectation is 6 straight to 10 not-straight. Since the sex-linked gen bow entered from the female, half the F2 males and females are bow. The half that are not-bow consist of 3 straight to 1 arc, so that both in the female classes and in the male classes there are 3 straight to 5 not-straight or in all 6 straight to 10 not-straight. The realized result, 248 straight to 307 not-straight, is more nearly a 3:4 ratio, due probably to a wrong classification of some of the bow as straight.
In this situation, the F2 expectation is 6 straight individuals to 10 not-straight. Since the sex-linked gene for bow came from the female, half of the F2 males and females have the bow trait. The other half that don't have bow consists of a ratio of 3 straight to 1 arc, meaning that in both the female and male groups, the ratio is 3 straight to 5 not-straight, or overall, 6 straight to 10 not-straight. The actual results show 248 straight individuals to 307 not-straight, which is closer to a 3:4 ratio, likely due to misclassification of some of the bow as straight.
LEMON BODY-COLOR.
Yellow body color.
(Plate I, figure 3.)
(__A_TAG_PLACEHOLDER_0__, figure 3.)
A few males of a new mutant with a lemon-colored body and wings appeared in August 1912. The lemon flies (Plate II, fig. 3) resemble quite closely the yellow flies (Plate II, fig. 4). They are paler and the bristles, instead of being brown, are black. These flies are so weak that despite most careful attention they get stuck to the food, so that they die before mating. The stock was at first maintained in mass from those cultures that gave the greatest percentage of lemon flies. In a few cases lemon males mated with their gray sisters left offspring, but the stock obtained in this way had still to be maintained by breeding heterozygotes, as stated above. But from the gray sisters heterozygous for lemon (bred to lemon males) some lemon females were also produced.
A few males of a new mutant with a lemon-colored body and wings appeared in August 1912. The lemon flies (Plate II, fig. 3) closely resemble the yellow flies (Plate II, fig. 4). They are paler, and their bristles are black instead of brown. These flies are so weak that even with the most careful attention, they get stuck to the food and die before mating. Initially, the stock was maintained in mass from those cultures that produced the highest percentage of lemon flies. In a few instances, lemon males mated with their gray sisters and produced offspring, but this stock still had to be maintained by breeding heterozygotes, as mentioned earlier. However, from the gray sisters heterozygous for lemon (bred to lemon males), some lemon females were also produced.
LINKAGE OF CHERRY, LEMON, AND VERMILION.
LINKAGE OF CHERRY, LEMON, AND VERMILION.
In order to study the linkage of lemon, the following experiment was carried out. Since it was impracticable to breed directly from the lemon flies, virgin females were taken from stock throwing lemon, and were mated singly to cherry vermilion males. Only a few of the females showed themselves heterozygous for lemon by producing lemon as well as gray sons. Half the daughters of such a pair are expected to be heterozygous for lemon and also for cherry and vermilion, which went in from the father. These daughters were mated singly to cherry vermilion males, and those that gave some lemon sons were continued, {49}and are recorded in table 24. The four classes of females were not separated from each other, but the total of females is given in the table.
To study the connection with lemon, the following experiment was conducted. Since it was impractical to breed directly from the lemon flies, virgin females were taken from the stock producing lemon and were paired one at a time with cherry vermilion males. Only a few of the females appeared to be heterozygous for lemon by producing both lemon and gray sons. Half of the daughters from such a pairing are expected to be heterozygous for lemon and also for cherry and vermilion, which they inherited from their father. These daughters were paired one at a time with cherry vermilion males, and those that had some lemon sons were continued, {49}and are recorded in table 24. The four classes of females were not separated, but the total number of females is provided in the table.
Table 24.—P1 lemon (het.) ♀ × cherry vermilion ♂ ♂. F1 wild-type ♀ × cherry vermilion ♂ ♂.
Table 24.—P1 lemon (het.) ♀ × cherry vermilion ♂ ♂. F1 wild-type ♀ × cherry vermilion ♂ ♂.
| Females. | 
| 
| 
| 
| Total ♂♂. | ||||
| Cherry ver- milion. | Lemon. | Cherry lemon. | Ver- milion. | Cherry | Lemon ver- milion. | Cherry lemon ver- milion. | Wild type. | ||
| 71 | 42 | 19 | 2 | 6 | 3 | 6 | 0 | 0 | 78 |
| 88 | 26 | 19 | 2 | 8 | 8 | 4 | 0 | 0 | 67 |
| 36 | 28 | 7 | 0 | 2 | 1 | 0 | 0 | 0 | 38 |
| 51 | 12 | 22 | 0 | 4 | 4 | 4 | 0 | 0 | 46 |
| 98 | 29 | 35 | 0 | 8 | 5 | 1 | 0 | 0 | 78 |
| 47 | 17 | 11 | 0 | 1 | 3 | 2 | 0 | 0 | 34 |
| 46 | 23 | 20 | 1 | 6 | 5 | 2 | 0 | 0 | 57 |
| 437 | 177 | 133 | 5 | 35 | 29 | 19 | 0 | 0 | 398 |
There are three loci involved in this cross, namely, cherry, lemon, and vermilion. Of these loci two were known, cherry and vermilion. The data are consistent with the assumption that the lemon locus is between cherry and vermilion, for the double cross-over classes (the smallest classes) are cherry lemon vermilion and wild type. The number of single cross-overs between cherry and lemon and between lemon and vermilion are also consistent with this assumption. Since lemon flies fail to emerge successfully, depending in part upon the condition of the bottle, the classes involving lemon are worthless in calculating crossing-over and are here ignored. In other words, lemon may be treated as though it did not appear at all, i. e., as a lethal. The not-lemon classes—cherry, vermilion, cherry vermilion, and wild type—give the following approximate cross-over values for the three loci involved: Cherry lemon, 15; lemon vermilion, 12; cherry vermilion, 27. The locus of lemon, calculated by interpolation, is at about 17.5.
There are three loci involved in this cross: cherry, lemon, and vermilion. Out of these, two were already known, cherry and vermilion. The data support the idea that the lemon locus is positioned between cherry and vermilion, since the double cross-over classes (the smallest groups) are cherry lemon vermilion and wild type. The number of single cross-overs between cherry and lemon and between lemon and vermilion also aligns with this idea. Because lemon flies don't successfully emerge, partly depending on the condition of the bottle, the classes involving lemon are not useful for calculating crossing-over and are disregarded here. In other words, lemon can be treated as if it didn't exist at all, i.e., as a lethal. The non-lemon classes—cherry, vermilion, cherry vermilion, and wild type—provide the following approximate cross-over values for the three loci involved: Cherry lemon, 15; lemon vermilion, 12; cherry vermilion, 27. The location of lemon, calculated through interpolation, is around 17.5.
LETHAL 2.
LETHAL 2.
In September 1912 a certain wild female produced 78 daughters and only 16 sons (Morgan, 1914b); 63 of these daughters were tested and 31 of them gave 2 females to 1 male, while 32 of them gave 1:1 sex-ratios. This shows that the mother of the original high sex-ratio was heterozygous for a recessive sex-linked lethal. In order to determine the position of this lethal, a lethal-bearing female was bred to an eosin (or white) miniature male, and those daughters that were heterozygous for eosin, lethal, and miniature were then back-crossed to {50}eosin miniature males. The daughters that result from such a cross give only the amount of crossing-over between eosin and miniature (as 29.7), but the males give the cross-over values for eosin lethal (9.9), lethal miniature (15.4), and eosin miniature (25.1). The data for this cross are given in table 25.
In September 1912, a particular wild female gave birth to 78 daughters and only 16 sons (Morgan, 1914b); 63 of these daughters were tested, and 31 of them produced 2 females for every 1 male, while 32 of them had a 1:1 sex ratio. This indicates that the mother with the unusually high sex ratio was heterozygous for a recessive sex-linked lethal. To determine the location of this lethal, a lethal-carrying female was bred with an eosin (or white) miniature male, and the daughters that were heterozygous for eosin, lethal, and miniature were then back-crossed to eosin miniature males. The daughters produced from such a cross reveal only the amount of recombination between eosin and miniature (which is 29.7), but the males provide the crossover values for eosin lethal (9.9), lethal miniature (15.4), and eosin miniature (25.1). The data for this cross are shown in table 25.
Table 25.—Total data upon linkage of eosin, lethal 2, and miniature, from Morgan, 1914b.
Table 25.—Total data on the connection of eosin, lethal 2, and miniature, from Morgan, 1914b.
| Females. | Males. | ||||||||
| Total. | Cross-overs. | Cross-over value. | 
| 
| 
| 
| Cross-over values. | ||
| Eosin lethal 2. | Lethal 2 miniature. | Eosin miniature. | |||||||
| 15,904 | 4,736 | 29.7 | 5,045 | 653 | 1,040 | 14 | 9.9 | 15.4 | 25.1 |
A similar experiment, in which eosin and vermilion were used instead of eosin and miniature, is summarized in table 26.
A similar experiment, where eosin and vermilion were used instead of eosin and miniature, is summarized in table 26.
Table 26.—Total data upon the linkage of eosin, lethal 2, and vermilion, from Morgan, 1914b.
Table 26.—Total data on the connection between eosin, lethal 2, and vermilion, from Morgan, 1914b.
| Females. | Males. | ||||||||
| Total. | Cross-overs. | Cross-over value. | 
| 
| 
| 
| Cross-over values. | ||
| Eosin lethal 2. | Lethal 2 vermilion. | Eosin vermilion. | |||||||
| 2,656 | 729 | 27.5 | 902 | 124 | 227 | 6 | 10.3 | 18.5 | 27.9 |
Considerable data in which lethal was not involved were also obtained in the course of these experiments and are included in the summary of the total data given in table 27.
Considerable data that did not involve lethal outcomes were also collected during these experiments and are included in the summary of the total data presented in table 27.
Table 27.—Summary of all data upon lethal 2, from Morgan, 1914b.
Table 27.—Summary of all data on lethal 2, from Morgan, 1914b.
| Gens. | Total. | Cross-overs. | Cross-over values. |
| White lethal 2 | 8,011 | 767 | 9.6 |
| White vermilion | 6,023 | 1,612 | 26.8 |
| White miniature | 36,021 | 11,048 | 30.7 |
| Lethal 2 vermilion | 1,400 | 248 | 17.7 |
| Lethal 2 miniature | 6,752 | 1,054 | 15.4 |
The amount of crossing-over between eosin and lethal is about 10 per cent and the amount of crossing-over between lethal and miniature is about 18 per cent. Since the amount of crossing-over between eosin {51}and miniature is over 30 per cent, the lethal factor must lie between eosin and miniature, somewhat nearer to eosin. It is impossible at present to locate lethal 2 accurately because of a real discrepancy in the data, which makes it appear that lethal 2 extends for a distance of about 5 units along the chromosome from about 10 to about 15. Work is being done which it is hoped will make clear the reason for this. For the present we may locate lethal 2 at the midpoint of its range, or at 12.5.
The crossover rate between eosin and lethal is about 10 percent, while the crossover rate between lethal and miniature is around 18 percent. Since the crossover rate between eosin {51} and miniature is over 30 percent, the lethal factor must be positioned between eosin and miniature, closer to eosin. Currently, it's challenging to pinpoint lethal 2 accurately due to a real discrepancy in the data, which suggests that lethal 2 spans about 5 units along the chromosome from approximately 10 to about 15. Ongoing research aims to clarify this issue. For now, we can place lethal 2 at the midpoint of its range, which is 12.5.
CHERRY.
CHERRY.
(Plate II, figure 9.)
(__A_TAG_PLACEHOLDER_0__, figure 9.)
The origin of the eye-color cherry has been given by Safir (Biol. Bull., 1913).
The origin of eye-color cherry was explained by Safir (Biol. Bull., 1913).
Cherry appeared (October 1912) in an experiment involving vermilion eye-color and miniature wings. This is the only time the mutant has ever come up, and although several of this mutant (males) appeared in Safir's experiment, they may have all come from the same mother. It is probable that the mutation occurred in the vermilion stock only a generation or so before the experiment was made, for otherwise cherry would be expected to be found also in the vermilion stock from which the mothers were taken; however, it was not found.
Cherry appeared (October 1912) in an experiment involving vermilion eye color and tiny wings. This is the only time this mutant has been observed, and although several of these mutants (males) showed up in Safir's experiment, they all may have come from the same mother. It's likely that the mutation occurred in the vermilion stock only a generation or so before the experiment took place, because otherwise, cherry would also be expected to be found in the vermilion stock from which the mothers were taken; however, it was not found.
A SYSTEM OF QUADRUPLE ALLELOMORPHS.
A system of four alleles.
Safir has described crosses between this eye-color and red, white, eosin, and vermilion. We conclude for reasons similar to those given by Morgan and Bridges (Jour. Exp. Zool., 1913) for the case of white and eosin, that cherry is an allelomorph of white and of eosin. This is not the interpretation followed in Safir's paper, where cherry is treated as though absolutely linked to white or to eosin. Both interpretations give, however, the same numerical result for each cross considered by itself. Safir's data and those which appear in this paper show that white, eosin, cherry, and a normal (red) allelomorph form a system of quadruple allelomorphs. If this interpretation is correct, then the linkage relations of cherry should be identical with those of white or of eosin.
Safir has described crosses between this eye color and red, white, eosin, and vermilion. We conclude, for reasons similar to those given by Morgan and Bridges (Jour. Exp. Zool., 1913) regarding white and eosin, that cherry is an allelomorph of both white and eosin. This differs from Safir's interpretation, where cherry is treated as though it’s completely linked to white or eosin. However, both interpretations yield the same numerical result for each cross considered individually. Safir's data, along with the information in this paper, show that white, eosin, cherry, and a normal (red) allelomorph form a system of quadruple allelomorphs. If this interpretation is accurate, then the linkage relationships of cherry should be the same as those of white or eosin.
LINKAGE OF CHERRY AND VERMILION.
LINKAGE OF CHERRY AND VERMILION.
The cross-over value for white (eosin) and vermilion, based on a very large amount of data, is about 31 units. An experiment of our own in which cherry was used with vermilion gave a cross-over value of 31 units, which is a close approximation to the cross-over value of white and vermilion. The cross which gave this data was that of a cherry vermilion (double recessive) male by wild females. The F1 wild-type flies inbred gave a single class of females (wild-type) and the males in four classes which show by the deviation from a 1:1:1:1 ratio the amount of crossing-over involved. {52}
The crossover value for white (eosin) and vermilion, based on a large amount of data, is about 31 units. In our own experiment using cherry with vermilion, we found a crossover value of 31 units, which closely matches the crossover value of white and vermilion. The cross for this data was between a cherry vermilion (double recessive) male and wild females. Inbreeding the F1 wild-type flies resulted in a single class of females (wild-type) and four classes of males, which revealed the amount of crossing-over that occurred through the deviation from a 1:1:1:1 ratio. {52}
In one of the F2 male classes of table 28 the simple eye-color cherry appeared for the first time (since the original mutant was vermilion as well as cherry). Safir has recorded a similar cross with like results.
In one of the F2 male classes of table 28, the simple eye color cherry showed up for the first time (since the original mutant was both vermilion and cherry). Safir has noted a similar cross with the same results.
Table 28.—P1 cherry vermilion ♂ ♂ × wild ♀ ♀. F1 wild-type ♀ ♀ × F1 wild-type ♂ ♂.
Table 28.—P1 cherry vermilion male × wild female. F1 wild-type female × F1 wild-type male.
| Reference. | Wild- type ♀♀. | Non-cross-over ♂. | Cross-over ♂. | Total ♂♂ | Cross- over value. | ||
| Cherry vermilion. | Wild- type. | Cherry. | Ver- milion. | ||||
| 160 C | 188 | 57 | 61 | 32 | 34 | 184 | 36 |
| 161 C | 256 | 85 | 93 | 40 | 52 | 270 | 34 |
| 162 C | 251 | 78 | 78 | 20 | 37 | 213 | 26 |
| 163 C | 229 | 76 | 95 | 34 | 33 | 238 | 28 |
| Total | 924 | 296 | 327 | 126 | 156 | 905 | 31 |
Some cherry males were bred to wild females. The F1 wild-type males and females inbred gave the results shown in table 29. Some of the cherry males thus produced were bred to their sisters. Cherry females as well as males resulted; and it was seen that the eye-color is the same in the males and females, in contradistinction to the allelomorph eosin, where there is a marked bicolorism (figs. 7, 8, Plate II). The cherry eye-color is almost identical with that of the eosin female, but is perhaps slightly more translucent and brighter.
Some cherry males were bred with wild females. The F1 wild-type males and females that were inbred produced the results shown in table 29. Some of the cherry males produced were bred with their sisters. Cherry females as well as males were produced; and it was noted that the eye color is the same in both males and females, unlike the allelomorph eosin, which shows a distinct bicolorism (figs. 7, 8, Plate II). The cherry eye color is almost the same as that of the eosin female, but it might be slightly more translucent and brighter.
Table 29.—P1 cherry ♂ ♂ × wild ♀ ♀. F1 wild-type ♀ ♀ × F1 wild-type ♂ ♂.
Table 29.—P1 cherry male × wild female. F1 wild-type female × F1 wild-type male.
| Reference. | Wild-type ♀. | Wild-type ♂. | Cherry ♂. |
| 15 I | 266 | 120 | 100 |
COMPOUNDS OF CHERRY.
CHERRY COMPOUNDS.
In order to examine the effect of the interaction of cherry and white in the same individual (i. e., white-cherry compound) cherry females were crossed to white males. This cross should give white-cherry females and cherry males. These white-cherry females were found (table 30) to be very much lighter than their brothers, the cherry males. The color of the pure cherry females and males is the same, but the substitution of one white for one cherry lowers the eye-color of the female below that of the cherry male. In eosin the white also lowers the eye-color of the compound female about in the same proportion as in the case of cherry. In the eosin the female starts at a higher degree of pigmentation than the male and dilution seems to bring her down {53}to the level of the male. But this coincidence of color between eosin male and white-eosin compound female is probably without significance, as shown by the results with cherry.
To explore how the combination of cherry and white works in the same individual (i.e., white-cherry hybrid), cherry females were crossed with white males. This cross should produce white-cherry females and cherry males. It was observed (see table 30) that the white-cherry females were significantly lighter than their brothers, the cherry males. The pure cherry males and females have the same color, but when one white is substituted for one cherry, it reduces the female's eye color compared to the cherry male. In eosin, the white also decreases the eye color of the hybrid female about the same as in the cherry case. In eosin, the female starts with a higher level of pigmentation than the male, and the dilution seems to reduce her pigmentation to match that of the male. However, this similarity in color between eosin males and white-eosin hybrid females probably doesn't mean much, as indicated by the cherry results.
Table 30.—P1 cherry ♀♀ × white ♂♂.
Table 30.—P1 cherry females × white males.
| Reference. | First generation. | |
| White-cherry compound ♀. | Cherry ♂. | |
| 9 M | 321 | 302 |
Eosin-cherry compound was also made. An eosin female was mated to a cherry male. The eosin-cherry daughters were darker than their eosin brothers. Inbred they gave the results shown in table 31.
Eosin-cherry compound was also created. An eosin female was paired with a cherry male. The eosin-cherry daughters were darker than their eosin brothers. When inbred, they produced the results shown in table 31.
Table 31.—P1 eosin ♀ × cherry ♂.
Table 31.—P1 eosin ♀ × cherry ♂.
| First generation. | Second generation. | ||||||
| Reference. | Eosin-cherry compound ♀♀. | Eosin ♂♂. | Reference. | Eosin and eosin-cherry compound ♀♀. | Cherry ♂. | Eosin ♂. | |
| 43C | 71 | 58 | 1I | 154 | 99 | 62 | |
| 2I | 174 | 74 | 77 | ||||
| 328 | 173 | 139 | |||||
Although in the F2 results there are two genotypic classes of females, namely, pure eosin and eosin-cherry compound, the eye-colors are so nearly the same that they can not be separated. The two classes of males can be readily distinguished; of these, one class, cherry, has the same color as the females, while the other class, eosin, is much lighter. Such an F2 group will perpetuate itself, giving one type of female (of three possible genotypic compositions, but somatically practically homogeneous) and two types of males, only one of which is like the females.
Although the F2 results show two genotypic classes of females, which are pure eosin and eosin-cherry compound, their eye colors are so similar that they can't be distinguished. The two classes of males can be easily identified; one class, cherry, has the same color as the females, while the other class, eosin, is much lighter. This F2 group will continue to reproduce itself, resulting in one type of female (with three possible genotypic combinations, but appearing nearly identical) and two types of males, only one of which resembles the females.
FUSED.
Fused.
In a cross between purple-eyed[6] males and black females there appeared in F2 (Nov. 4, 1912) a male having the veins of the wing arranged as shown in text-figure D b. It will be seen that the third and the fourth longitudinal veins are fused from the base to and beyond the {54}point at which in normal flies the anterior cross-vein lies. The cross-vein and the cell normally cut off by it are absent. There are a number of other features (see fig. D c) characteristic of this mutation: the wings are held out at a wide angle from the body, the ocelli are very much reduced in size or entirely absent, the bristles around the ocelli are usually small. The females are absolutely sterile, not only with their own, but with any males.
In a cross between purple-eyed[6] males and black females, a male was observed in F2 (Nov. 4, 1912) that had the wing veins arranged as shown in text-figure D b. You can see that the third and fourth longitudinal veins are fused from the base and extend beyond the point where the anterior cross-vein is located in typical flies. The cross-vein and the cell that it usually separates are missing. There are several other features (see fig. D c) that are characteristic of this mutation: the wings are positioned at a wide angle from the body, the ocelli are significantly reduced in size or completely absent, and the bristles around the ocelli are generally small. The females are completely sterile, unable to breed not only with their own kind but also with any males.
Fused males by wild females gave wild-type males and females. Inbred these gave the results shown in table 32. The fused character reappeared only in the F2 males, showing that it is a recessive sex-linked character.
Fused males from wild females produced wild-type males and females. When inbred, these resulted in the outcomes shown in table 32. The fused trait appeared again only in the F2 males, indicating that it is a recessive sex-linked trait.
Table 32.—P1 fused ♂ × wild ♀♀.
Table 32.—P1 fused male × wild females.
| First generation. | Second generation. | ||||||
| Reference. | Wild-type ♀♀. | Wild-type ♂♂. | Reference. | Wild-type ♀♀. | Wild-type ♂♂. | Fused ♂♂. | |
| 4I | 66 | 43 | 190C | 258 | 96 | 115 | |
| 14I | 239 | 105 | 90 | ||||
| Total | 497 | 201 | 205 | ||||
The reciprocal cross was tried many times, but is impossible, owing to the sterility of the females. Since the fused females are sterile to fused males, the stock is kept up by breeding heterozygous females to fused males.
The reciprocal cross was attempted many times, but it’s impossible due to the sterility of the females. Since the fused females are sterile with the fused males, the population is maintained by breeding heterozygous females with the fused males.
By means of the following experiments the position of fused in the X chromosome was determined. A preliminary test was made by mating with eosin, whose factor lies near the left end of the X chromosome series.
By conducting the following experiments, the location of fused on the X chromosome was identified. A preliminary test was performed by mating with eosin, whose factor is located close to the left end of the X chromosome series.
LINKAGE OF EOSIN AND FUSED.
LINKAGE OF EOSIN AND FUSED.
Fused (red-eyed) males mated to eosin (not-fused) females gave wild-type daughters and eosin sons, which inbred gave the classes shown in table 33.
Fused (red-eyed) males paired with eosin (not-fused) females produced wild-type daughters and eosin sons, which, when inbred, resulted in the classes shown in table 33.
Table 33.—P1 eosin ♀♀ × fused ♂♂. F1 wild-type ♀♀ × F1 eosin ♂♂.
Table 33.—P1 eosin ♀♀ × fused ♂♂. F1 wild-type ♀♀ × F1 eosin ♂♂.
| Reference. | Females. | Non-cross-over ♂♂. | Cross-over ♂♂. | Total males. | Cross- over value. | ||
| Eosin. | Fused. | Eosin fused. | Wild- type. | ||||
| 56I | 496 | 131 | 113 | 82 | 104 | 430 | 43 |
The data give 43 per cent of crossing-over, which places fused far to the right or to the left of eosin. The latter position is improbable, since eosin already lies very near the extreme left end of the known series. Therefore, since 43 per cent would place the factor nearly at the right end of the series, the next step was to test its relation to a factor like bar that lies at the right end of the chromosome. By mating to bar alone we could only get the linkage to bar without discovering on which side of bar the new factor lies, but by mating to a fly that carries still another sex-linked factor, known to lie to the left of bar, the information gained should show the relative order of the factors involved. Furthermore, since, by making a back-cross, both males and females give the same kind of data (and need not be separated), the experiment was made in this way. In order to have material for such an experiment double mutant stocks of vermilion fused and also of bar fused were made up.
The data shows 43 percent of crossing-over, which positions fused far to the right or to the left of eosin. The latter placement is unlikely, since eosin is already very close to the extreme left end of the known series. Therefore, because 43 percent would place the factor nearly at the right end of the series, the next step was to test its relationship to a factor like bar that is located at the right end of the chromosome. By mating with bar alone, we could only identify the linkage to bar without determining which side of bar the new factor is on. However, by mating with a fly that has another sex-linked factor, known to be to the left of bar, we should be able to discern the relative order of the factors involved. Additionally, since both males and females provide the same type of data in a back-cross (and don’t need to be separated), the experiment was conducted this way. To have material for such an experiment, double mutant stocks of vermilion fused and bar fused were prepared.

Fig. D.—a, normal wing; b and c, fused wings. c shows a typical fused wing. The most striking feature is the closure of the cell between the third and fourth longitudinal veins with the elimination of the cross-vein; the veins at the base of the wing differ from those in the normal shown in a. b shows the normal position in which the fused wings are held. The fusion of the veins in b is unusually complete.
Fig. D.—a, normal wing; b and c, fused wings. c displays a typical fused wing. The most noticeable feature is the closure of the cell between the third and fourth longitudinal veins, along with the removal of the cross-vein; the veins at the base of the wing are different from those in the normal wing shown in a. b illustrates the usual position in which the fused wings are held. The fusion of the veins in b is remarkably complete.
LINKAGE OF VERMILION, BAR, AND FUSED.
LINKAGE OF VERMILION, BAR, AND FUSED.
Males from the stock of (red) bar fused were mated to vermilion (not-bar, not-fused) females, and produced bar females and vermilion males. The bar F1 daughters were back-crossed to vermilion fused males and produced the classes of offspring shown in table 34.
Males from the (red) bar fused line were paired with vermilion (not-bar, not-fused) females, resulting in bar females and vermilion males. The bar F1 daughters were back-crossed to vermilion fused males, producing the classes of offspring shown in table 34.
Table 34.—P1 vermilion ♀ ♀ × bar fused ♂ ♂. B. C. F1 bar ♀ × vermilion fused ♂ ♂.
Table 34.—P1 vermilion female × bar fused male. B. C. F1 bar female × vermilion fused male.
| Reference. | 
| 
| 
| 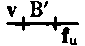
| Total. | Cross-over values. | ||||||
| Ver- milion. | Bar fused. | Ver- milion bar fused. | Wild- type. | Ver- milion fused. | Bar. | Ver- milion bar. | Fused. | Ver- milion bar. | Bar fused. | Ver- milion fused. | ||
| 140 I | 137 | 130 | 35 | 40 | 5 | 8 | .. | .. | 355 | 21 | 4 | 25 |
| 141 I | 144 | 137 | 38 | 41 | 4 | 2 | .. | .. | 366 | 22 | 2 | 23 |
| 142 I | 153 | 120 | 43 | 58 | 6 | 7 | 1 | .. | 388 | 26 | 4 | 29 |
| 143 I | 153 | 92 | 44 | 41 | 3 | 7 | 3 | 1 | 344 | 26 | 4 | 28 |
| 145 I | 69 | 62 | 29 | 19 | 1 | .. | 1 | .. | 181 | 27 | 1 | 27 |
| 146 I | 96 | 103 | 30 | 34 | 7 | 3 | .. | .. | 273 | 23 | 4 | 26 |
| 156 I | 62 | 45 | 25 | 27 | 1 | 4 | .. | .. | 164 | 32 | 3 | 35 |
| 157 I | 93 | 57 | 11 | 31 | 2 | 2 | .. | 2 | 198 | 22 | 3 | 23 |
| Total. | 907 | 746 | 255 | 291 | 29 | 33 | 5 | 3 | 2,269 | 24 | 3 | 27 |
The data show that the factor for fused lies about 3 units to the right of bar. This is the furthest point yet obtained to the right. The reasons for locating fused to the right of bar are that, if it occupies such a position, then the double cross-over classes (which are expected to be the smallest classes) should be vermilion bar and fused, and these are, in fact, the smallest classes. The order of factors is, then, vermilion, bar, fused. This order is confirmed by the result that the number of cross-overs between fused and vermilion is greater than that between bar and vermilion.
The data indicate that the factor for fused is positioned about 3 units to the right of bar. This is the farthest point reached to the right so far. The reasons for placing fused to the right of bar are that, if it takes that position, then the double crossover classes (which are expected to be the smallest classes) should be vermilion bar and fused, and these are indeed the smallest classes. The order of factors is, therefore, vermilion, bar, fused. This order is supported by the finding that the number of crossovers between fused and vermilion is greater than that between bar and vermilion.
In order to obtain data to balance viability effects, the following experiment was made:
In order to gather data to balance viability effects, the following experiment was conducted:
Vermilion (not-bar) fused males were bred to (red) bar (not-fused) females. The daughters and sons were bar. The daughters were back-crossed, singly, to vermilion fused males and gave the results shown in table 35. Each female was also transferred to a second culture bottle, so that for each female there are two broods given consecutively (82, 82′, etc.) in table 35.
Vermilion (not-bar) fused males were bred with (red) bar (not-fused) females. The offspring were bar. The daughters were back-crossed, one at a time, to vermilion fused males and produced the results shown in table 35. Each female was also moved to a second culture bottle, so that for each female there are two broods listed consecutively (82, 82′, etc.) in table 35.
Table 35.—P1 bar ♀ ♀ × vermilion fused ♂ ♂. B. C. F1 bar ♀ × vermilion fused ♂ ♂.
Table 35.—P1 bar ♀ ♀ × vermilion fused ♂ ♂. B. C. F1 bar ♀ × vermilion fused ♂ ♂.
| Reference. | 
| 
| 
| 
| Total. | Cross-over values. | ||||||
| Ver- milion fused. | Bar. | Ver- milion bar. | Fused. | Ver- milion. | Bar fused. | Ver- milion bar fused. | Wild- type. | Ver- milion bar. | Bar fused. | Ver- milion fused. | ||
| 82 | 165 | 165 | 63 | 57 | 8 | 7 | 1 | .. | 466 | 26 | 3 | 29 |
| 82′ | 104 | 87 | 26 | 24 | .. | 4 | .. | .. | 245 | 20 | 2 | 22 |
| 83 | 128 | 164 | 51 | 39 | 6 | 4 | .. | .. | 392 | 23 | 3 | 26 |
| 83′ | 100 | 94 | 28 | 30 | 4 | 4 | .. | .. | 260 | 22 | 3 | 25 |
| 89 | 85 | 105 | 23 | 24 | 5 | 2 | .. | .. | 244 | 19 | 3 | 22 |
| 89′ | 78 | 91 | 21 | 27 | 1 | 2 | .. | 1 | 221 | 22 | 2 | 23 |
| 90 | 86 | 85 | 30 | 28 | 5 | .. | .. | .. | 234 | 25 | 2 | 27 |
| 90′ | 33 | 38 | 22 | 14 | 4 | 1 | .. | 1 | 113 | 33 | 5 | 36 |
| 91 | 125 | 107 | 41 | 31 | 1 | 1 | .. | .. | 306 | 24 | 1 | 24 |
| 91′ | 91 | 95 | 31 | 25 | 5 | 1 | .. | 2 | 250 | 23 | 3 | 25 |
| 92 | 109 | 136 | 41 | 24 | 4 | 2 | .. | .. | 316 | 21 | 2 | 23 |
| 92′ | 100 | 105 | 29 | 29 | .. | 1 | .. | 1 | 265 | 22 | 1 | 22 |
| 93 | 75 | 67 | 19 | 20 | .. | 1 | .. | .. | 182 | 21 | 1 | 22 |
| 93′ | 68 | 94 | 31 | 17 | 1 | 1 | .. | .. | 212 | 23 | 1 | 24 |
| 94 | 84 | 96 | 31 | 35 | 8 | 1 | .. | .. | 255 | 26 | 4 | 29 |
| 94′ | 61 | 73 | 20 | 22 | 5 | 4 | .. | .. | 185 | 23 | 5 | 28 |
| 95 | 84 | 102 | 27 | 26 | 3 | 3 | .. | .. | 245 | 22 | 2 | 24 |
| 96 | 144 | 148 | 43 | 34 | 1 | 2 | .. | 1 | 373 | 21 | 1 | 21 |
| 97 | 81 | 96 | 25 | 20 | 5 | 3 | .. | .. | 230 | 20 | 4 | 23 |
| 98 | 107 | 112 | 39 | 33 | 1 | 2 | .. | .. | 294 | 25 | 1 | 26 |
| Firsts | 1,273 | 1,383 | 433 | 371 | 47 | 28 | 1 | 1 | 3,537 | 23 | 2 | 25 |
| Seconds | 635 | 677 | 208 | 188 | 20 | 18 | .. | 5 | 1,751 | 23 | 3 | 25 |
| Total. | 1,908 | 2,060 | 641 | 559 | 67 | 46 | 1 | 6 | 5,288 | 23 | 2.3 | 25 |
Table 36.—Linkage of vermilion, bar, and fused with balanced viability.
Table 36.—Connection of vermilion, bar, and fused with balanced viability.

| 
| 
| 
| Total. | |
| 5,621 | 1,756 | 175 | 15 | 7,567 | |
| Percentage | 74.3 | 23.19 | 2.31 | 0.2 |
Some additional data bearing on the linkage of vermilion and fused were obtained. Males of (red) fused stock were bred to vermilion (not-fused) females, and gave wild-type females and vermilion males, which inbred gave the results shown in table 37.
Some additional data related to the connection between vermilion and fused were gathered. Males of (red) fused stock were bred with vermilion (not-fused) females, resulting in wild-type females and vermilion males, which when inbred produced the results shown in table 37.
The percentage of cross-overs between vermilion and fused is here 27, which is in agreement with the 26 per cent of the preceding experiment.
The percentage of crossovers between vermilion and fused is 27% here, which matches the 26% from the previous experiment.
The converse experiment, namely, red (not-fused) females by vermilion fused males also gave, when the wild-type daughters were {58}back-crossed to vermilion fused males, a linkage value of 27 units. Two 10-day broods were reared from each female. The data given in table 38 show that the percentage of crossing-over does not change as the flies get older. The locus of fused on the basis of all of the data is at 59.5.
The reverse experiment, where non-fused red females were paired with fused vermilion males, also showed that when the wild-type daughters were back-crossed with fused vermilion males, there was a linkage value of 27 units. Two broods, each lasting 10 days, were raised from each female. The data in table 38 indicate that the percentage of crossing-over remains consistent as the flies age. Based on all the data, the location of the fused gene is at 59.5.
Table 37.—P1 vermilion ♀ ♀ × fused ♂ ♂. F1 wild-type ♀ ♀ × F1 vermilion ♂ ♂.
Table 37.—P1 vermilion female × fused male. F1 wild-type female × F1 vermilion male.
| Reference. | Females. | Non-cross-over ♂ ♂. | Cross-over ♂ ♂. | Total ♂ ♂. | Cross- over values. | ||
| Vermilion. | Fused. | Vermilion fused. | Wild- type. | ||||
| 79 I | 299 | 93 | 96 | 37 | 36 | 262 | 28 |
| 80 I | 245 | 93 | 60 | 28 | 27 | 208 | 26 |
| 81 I | 263 | 101 | 63 | 22 | 40 | 226 | 27 |
| Total. | 807 | 287 | 219 | 87 | 103 | 696 | 27 |
Table 38.—P1 wild ♀ ♀ × vermilion fused ♂ ♂. F1 wild-type ♀ × F1 wild-type ♂ ♂.
Table 38.—P1 wild females × vermilion fused males. F1 wild-type females × F1 wild-type males.
| Reference. | Females. | Non-cross-over ♂ ♂. | Cross-over ♂ ♂. | Total ♂ ♂. | Cross- over values. | ||
| Vermilion fused. | Wild- type. | Vermilion. | Fused. | ||||
| 52 | 96 | 25 | 30 | 16 | 11 | 82 | 33 |
| 52′ | 176 | 59 | 64 | 24 | 19 | 166 | 26 |
| 53 | 60 | 20 | 22 | 9 | 6 | 57 | 26 |
| 53′ | 76 | 21 | 27 | 11 | 10 | 69 | 31 |
| 54 | 88 | 35 | 38 | 14 | 16 | 103 | 29 |
| 54′ | 60 | 22 | 20 | 8 | 9 | 59 | 29 |
| 57 | 61 | 22 | 20 | 7 | 11 | 60 | 30 |
| 57′ | 170 | 47 | 54 | 24 | 19 | 144 | 30 |
| 58 | 128 | 37 | 55 | 14 | 10 | 116 | 21 |
| 58′ | 144 | 38 | 64 | 16 | 15 | 133 | 23 |
| Firsts | 433 | 139 | 165 | 60 | 54 | 418 | 27 |
| Seconds | 626 | 187 | 229 | 83 | 72 | 571 | 27 |
| Total | 1,059 | 326 | 394 | 143 | 126 | 989 | 27 |
FORKED.
Split.
On November 19, 1912 there appeared in a stock of a double recessive eye-color, vermilion maroon, a few males which showed a novel form of the large bristles (macrochætæ) upon the head and thorax. In this mutation (text-fig. E) the first of several which affect the shape and distribution of the bristles, the macrochætæ, instead of {59}being long, slender, and tapered (see Plate 1, fig. I), are greatly shortened and crinkled as though scorched. The ends are forked or branched, bent sharply, or merely thickened. The bristles which are most distorted are those upon the scutellum, where they are sometimes curled together into balls.
On November 19, 1912, a group of males with a double recessive eye color, vermilion maroon, revealed a new form of large bristles (macrochætæ) on their heads and thoraxes. In this mutation (text-fig. E), which is the first of several that change the shape and distribution of the bristles, the macrochætæ, instead of being long, slender, and tapered (see Plate 1, fig. I), are significantly shortened and crinkled as if they’ve been scorched. The ends are forked or branched, sharply bent, or just thickened. The most distorted bristles are those on the scutellum, where they sometimes curl together into balls.
LINKAGE OF VERMILION AND FORKED.
LINKAGE OF VERMILION AND FORKED.
 Fig. E.—Forked bristles.
Fig. E.—Forked bristles.
Since forked arose in vermilion stock, the double recessive for these two sex-linked factors could be used in testing the linkage relations of the mutation. Vermilion forked males were crossed to wild females and gave wild-type males and females, which inbred gave in F2 the results shown in table 39. Forked reappeared only in the males in the following proportion: not-forked ♀, 742; not-forked ♂, 346; forked ♂, 301. The result shows that the character is a sex-linked recessive.
Since forked originated in the vermilion stock, the double recessive for these two sex-linked factors could be used to test the linkage relations of the mutation. Vermilion forked males were crossed with wild females, producing wild-type males and females, which when inbred resulted in the F2 outcomes shown in table 39. Forked appeared only in the males in the following proportions: not-forked ♀, 742; not-forked ♂, 346; forked ♂, 301. The results indicate that the trait is a sex-linked recessive.
Table 39.—P1 wild ♀ ♀ × vermilion-forked ♂ ♂. F1 wild-type ♀ ♀ × F1 wild-type ♂ ♂.
Table 39.—P1 wild females × vermilion-forked males. F1 wild-type females × F1 wild-type males.
| Reference. | Wild-type ♀♀. | Non-cross-over ♂ ♂. | Cross-over ♂ ♂. | Total ♂ ♂. | Cross- over values. | ||
| Vermilion forked. | Wild- type. | Vermilion. | Forked. | ||||
| 9 I | 366 | 113 | 123 | 49 | 41 | 326 | 28 |
| 11 I | 376 | 116 | 150 | 42 | 31 | 339 | 22 |
| Total. | 742 | 229 | 273 | 91 | 72 | 665 | 25 |
In table 39 vermilion forked and wild-type are non-cross-overs, and vermilion and forked are cross-overs, giving a cross-over value of 25 units. The locus, therefore, is 25 units to the right or to the left of vermilion, that is, either about 58 or 8 units from the yellow locus.
In table 39, vermilion forked and wild-type are non-cross-overs, while vermilion and forked are cross-overs, leading to a cross-over value of 25 units. This means the locus is 25 units to the right or left of vermilion, specifically around 58 or 8 units from the yellow locus.
LINKAGE OF CHERRY AND FORKED.
LINKAGE OF CHERRY AND FORKED.
Forked males were crossed to cherry females (cherry has the same locus as white, which is about 1 unit from yellow) and gave wild-type females and cherry males. These gave in F2 the results shown in table 40. The non-cross-overs (cherry and forked) plus the cross-overs (cherry forked and wild type) divided into the cross-overs give a cross-over value of 46 units, which shows that the locus lies to the right of vermilion, because if it had been to the left, the value would have been 8 (i. e., 33-25) instead of 33+25=58. The difference between 58 {60}and 46 is due to the expected amount of double crossing-over. In fact, for a distance as long as 58 an almost independent behavior of linked gens is to be expected.
Forked males were bred with cherry females (cherry shares the same locus as white, which is about 1 unit away from yellow) and produced wild-type females and cherry males. These results in F2 are shown in table 40. The non-crossovers (cherry and forked) along with the crossovers (cherry forked and wild type) split into the crossovers to give a crossover value of 46 units, indicating that the locus is located to the right of vermilion. If it had been to the left, the value would have been 8 (i.e., 33-25) instead of 33+25=58. The difference between 58 {60} and 46 is due to the expected amount of double crossing-over. In fact, for a distance as long as 58, almost independent behavior of linked genes is expected.
Table 40.—P1 cherry ♀ ♀ × forked ♂ ♂. F1 wild-type ♀ ♀ × F1 cherry ♂ ♂.
Table 40.—P1 cherry female × forked male. F1 wild-type female × F1 cherry male.
| Reference. | Females. | Non-cross-over ♂ ♂. | Cross-over ♂ ♂. | Total ♂ ♂. | Cross- over values. | |||
| Cherry. | Wild- type. | Cherry. | Forked. | Cherry forked. | Wild- type. | |||
| 25 | 129 | 145 | 73 | 70 | 65 | 68 | 276 | 48 |
| 25′ | 167 | 148 | 74 | 82 | 66 | 88 | 310 | 50 |
| 36 | 96 | 88 | 52 | 52 | 35 | 51 | 190 | 45 |
| 36′ | 57 | 76 | 41 | 32 | 24 | 30 | 127 | 43 |
| 84 | 76 | 86 | 40 | 34 | 38 | 26 | 138 | 46 |
| 84′ | 62 | 71 | 24 | 39 | 25 | 28 | 116 | 46 |
| 85 | 114 | 86 | 43 | 78 | 41 | 53 | 215 | 44 |
| 85′ | 98 | 95 | 48 | 63 | 52 | 46 | 209 | 47 |
| 86 | 307 | 323 | 152 | 144 | 118 | 165 | 579 | 49 |
| 87 | 351 | 341 | 183 | 213 | 160 | 147 | 703 | 45 |
| 88 | 244 | 246 | 142 | 142 | 107 | 104 | 495 | 43 |
| Total. | 1,701 | 1,705 | 872 | 949 | 731 | 806 | 3,358 | 46 |
LINKAGE OF FORKED, BAR, AND FUSED.
LINKAGE OF FORKED, BAR, AND FUSED.
This value of 58 gave the furthest locus to the right obtained up to that time, since forked is slightly beyond rudimentary. Later, the locus for bar-eye was found still farther to the right, and the locus for fused even farther to the right than bar. A cross was made involving these three gens. A forked (not-bar) fused male was bred to a (not-forked) bar (not-fused) female and gave bar females and males. The F1 females were back-crossed singly to forked fused males with the result shown in table 41.
This value of 58 provided the farthest position to the right achieved up to that point, since forked is slightly beyond basic. Later, the position for bar-eye was discovered even further to the right, and the position for fused was found to be even farther to the right than bar. A cross was made involving these three genes. A forked (not-bar) fused male was bred with a (not-forked) bar (not-fused) female, resulting in bar females and males. The F1 females were back-crossed individually with forked fused males, leading to the results shown in table 41.
Table 41.—P1 bar ♀ ♀ × forked fused ♂ ♂. B. C. F1 bar ♀ × forked fused ♂ ♂.
Table 41.—P1 bar female × forked fused male. B. C. F1 bar female × forked fused male.
| Reference. | 
| 
| 
| 
| Total. | ||||
| Forked fused. | Bar. | Forked bar. | Fused. | Forked. | Bar fused. | Forked bar fused. | Wild- type. | ||
| 163 | 45 | 55 | .. | 1 | 4 | 2 | .. | .. | 108 |
| 164 | 71 | 90 | .. | .. | 4 | 1 | .. | .. | 166 |
| 165 | 97 | 106 | .. | .. | 2 | 4 | .. | .. | 209 |
| 11 | 21 | 35 | .. | .. | 1 | 2 | .. | .. | 59 |
| 33 | 15 | 23 | .. | .. | .. | 1 | .. | .. | 39 |
| Total. | 250 | 309 | .. | 1 | 11 | 10 | .. | .. | 581 |
The same three points were combined in a different way, namely, by mating forked females to bar fused males. The bar daughters were back-crossed to forked fused males and gave the results shown in table 42.
The same three points were combined differently by mating forked females with bar fused males. The bar daughters were back-crossed to forked fused males, producing the results shown in table 42.
Table 42.—P1 forked ♀ ♀ × bar fused ♂ ♂. B.C. F1 bar ♀ × forked fused ♂ ♂.
Table 42.—P1 forked female female × bar fused male male. B.C. F1 bar female × forked fused male male.
| Reference. | 
| 
| 
| 
| Total. | ||||
| Forked. | Fused bar. | Forked bar fused. | Wild- type. | Forked fused. | Bar. | Forked bar. | Fused. | ||
| 158 | 131 | 124 | 1 | .. | 3 | 3 | .. | .. | 262 |
| 159 | 31 | 45 | .. | .. | .. | .. | .. | .. | 76 |
| 160 | 29 | 23 | .. | .. | 1 | 2 | .. | .. | 55 |
| 161 | 24 | 11 | 1 | .. | .. | .. | .. | .. | 36 |
| 162 | 96 | 91 | 2 | .. | 1 | 1 | .. | .. | 191 |
| Total. | 311 | 294 | 4 | .. | 5 | 6 | .. | .. | 620 |
By combining the results of tables 41 and 42 data are obtained for cross-over values from which (by balancing the inviable classes, as explained in table 43) the element of inviability is reduced to a minimum.
By combining the results of tables 41 and 42, data is collected for cross-over values, from which (by balancing the non-viable classes, as explained in table 43) the factor of non-viability is minimized.
Table 43.
Table 43.
|
|
|
|
| Total. | |
| 1,164 | 5 | 32 | 0 | 1,201 | |
| Per cent. | 96.9 | 0.42 | 2.7 | 0 |
The linkages involved in these data are very strong. The cross-overs between forked and bar number only 5 in a total of 1,201, which gives less than 0.5 per cent of crossing-over. There are 32 cross-overs or 2.7 per cent between bar and fused. The value for forked fused is the sum of the two other values, or 3.1 per cent.
The connections in this data are very strong. The crossovers between forked and bar are only 5 out of a total of 1,201, which is less than 0.5 percent of crossover. There are 32 crossovers, or 2.7 percent, between bar and fused. The value for forked fused is the sum of the other two values, which is 3.1 percent.
LINKAGE OF SABLE, RUDIMENTARY, AND FORKED.
LINKAGE OF SABLE, BASIC, AND FORKED.
Rudimentary, forked, bar, and fused form a rather compact group at the right end of the chromosome, as do yellow, lethal 1, white, abnormal, etc., at the zero end. The following two experiments were made to determine more accurately the interval between rudimentary and the other members of this group. A sable rudimentary forked {62}male mated to a wild female gave wild-type sons and daughters. These inbred give the results shown in table 44.
Rudimentary, forked, bar, and fused form a tight cluster at the right end of the chromosome, just like yellow, lethal 1, white, abnormal, and others do at the left end. Two experiments were conducted to more precisely measure the distance between rudimentary and the other members of this group. A sable rudimentary forked {62} male mated with a wild female produced wild-type sons and daughters. These inbred offspring produced the results shown in table 44.
Table 44.—P1 sable rudimentary forked ♂ × wild ♀. F1 wild-type ♀ × F1 wild-type ♂ ♂.
Table 44.—P1 black basic forked male × wild female. F1 wild-type female × F1 wild-type male male.
| Reference. | Wild- type ♀♀. | 
| 
| 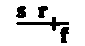
|  | ||||
| Sable rudi- mentary forked. | Wild- type. | Sable. | Rudi- mentary forked. | Sable rudi- mentary. | Forked. | Sable forked. | Rudi- mentary. | ||
| 264 | 98 | 28 | 17 | 2 | 5 | 1 | 1 | .. | .. |
| 265 | 97 | 29 | 54 | 4 | 9 | .. | .. | .. | .. |
| 266 | 114 | 42 | 49 | 11 | 11 | .. | 2 | .. | .. |
| Total | 309 | 99 | 120 | 17 | 25 | 1 | 3 | .. | .. |
There were 265 males, of which 42 were cross-overs between sable and rudimentary and 4 between rudimentary and forked. The values found are: sable rudimentary, 16; rudimentary forked, 1.5; sable forked, 17.
There were 265 males, including 42 that were hybrids between sable and rudimentary and 4 between rudimentary and forked. The values recorded are: sable rudimentary, 16; rudimentary forked, 1.5; sable forked, 17.
LINKAGE OF RUDIMENTARY, FORKED, AND BAR.
LINKAGE OF BASIC, FORKED, AND BAR.
The three gens, rudimentary, forked, and bar, form a very compact group. A rudimentary forked male was crossed to bar females and the daughters (bar) were back-crossed singly to rudimentary forked males, the results being shown in table 45.
The three gens—rudimentary, forked, and bar—create a very tight-knit group. A rudimentary forked male was mated with bar females, and the daughters (bar) were back-crossed one at a time to rudimentary forked males, with the results displayed in table 45.
Table 45.—P1 rudimentary forked ♂ × bar ♀. B.C. F1 bar ♀ × rudimentary forked ♂ ♂.
Table 45.—P1 rudimentary forked male × bar female. B.C. F1 bar female × rudimentary forked male male.
| Reference. | 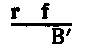
| 
| 
|  | ||||
| Rudi- mentary forked. | Bar. | Rudi- mentary bar. | Forked. | Rudi- mentary forked bar. | Wild- type. | Rudi- mentary. | Forked bar. | |
| 267 | 56 | 104 | .. | 2 | 1 | 1 | .. | .. |
| 268 | 82 | 86 | 1 | 2 | .. | .. | .. | .. |
| 269 | 68 | 101 | .. | .. | .. | 1 | .. | .. |
| Total | 206 | 291 | 1 | 4 | 1 | 2 | .. | .. |
SHIFTED.
SHIFTE⟩D.
Shifted appeared (January 1913) in a stock culture of vermilion dot. The chief characteristic of this mutant is that the third longitudinal vein (see text-fig. F) does not reach the margin as it does in the normal fly. The vein is displaced toward the fourth throughout its length, and only very rarely does it extend far enough to join the marginal vein. The cross-vein between the third and the fourth veins is often absent because of the shifting. The flies themselves are smaller than normal. The wings are held out from the body at a wide angle. The two posterior bristles of the scutellum are much reduced in size and stick straight up—a useful landmark by which just-hatched shifted flies may be recognized, even though the wings are not expanded.
Shifted appeared (January 1913) in a stock culture of vermilion dot. The main feature of this mutant is that the third longitudinal vein (see text-fig. F) doesn't reach the edge like it does in the normal fly. The vein is pushed towards the fourth vein along its entire length, and only very rarely does it extend far enough to connect with the marginal vein. The cross-vein between the third and fourth veins is often missing due to this shift. The flies themselves are smaller than usual. Their wings are held out from the body at a wide angle. The two back bristles of the scutellum are much smaller and stand straight up—a handy marker to identify freshly hatched shifted flies, even when their wings aren't spread.
LINKAGE OF SHIFTED AND VERMILION.
LINKAGE OF SHIFTED AND RED.
Since shifted arose in vermilion, the double recessive shifted vermilion was available for the following linkage experiment: shifted vermilion males by wild females gave wild-type males and females which inbred gave the data shown in table 46.
Since shifted appeared in vermilion, the double recessive shifted vermilion was available for the following linkage experiment: shifted vermilion males crossed with wild females produced wild-type males and females, which when inbred, yielded the data shown in table 46.

Fig. F.—Shifted venation. The third longitudinal vein is shifted toward the fourth and fails to reach the margin. Cross-vein between third and fourth longitudinal veins is lacking.
Fig. F.—Shifted venation. The third longitudinal vein is moved towards the fourth and doesn't reach the edge. The cross-vein between the third and fourth longitudinal veins is missing.
Disregarding the eye-color, the following is a summary of the preceding results: wild-type ♀, 1,001; wild-type ♂, 437; shifted ♂, 328. The result shows that shifted is a sex-linked recessive. The data of table 46 show that the locus of shifted lies about 15 units on one side or the other of vermilion, which from the calculated position of vermilion at 33 would give a position for shifted at either 18 or 48 from yellow.
Disregarding eye color, here's a summary of the previous results: wild-type female, 1,001; wild-type male, 437; shifted male, 328. This indicates that shifted is a sex-linked recessive trait. The data in table 46 show that the locus of shifted is about 15 units on either side of vermilion, which, given the calculated position of vermilion at 33, places shifted at either 18 or 48 from yellow.
Table 46.—P1 shifted vermilion ♂ ♂ × wild ♀ ♀. F1 wild-type ♀ × F1 wild-type ♂ ♂.
Table 46.—P1 shifted vermilion ♂ ♂ × wild ♀ ♀. F1 wild-type ♀ × F1 wild-type ♂ ♂.
| Reference. | Females. | Non-cross-over ♂ ♂. | Cross-over ♂ ♂. | Total ♂ ♂. | Cross- over values. | ||
| Shifted vermilion. | Wild- type. | Shifted. | Vermilion. | ||||
| 13 | 345 | 79 | 115 | 8 | 25 | 227 | 15 |
| 29 | 68 | 20 | 32 | 3 | 4 | 59 | 12 |
| 30 | 191 | 37 | 54 | 5 | 13 | 109 | 17 |
| 31 | 151 | 41 | 65 | 17 | 13 | 136 | 22 |
| 33 | 133 | 49 | 40 | 4 | 6 | 99 | 10 |
| 34 | 113 | 56 | 59 | 9 | 11 | 135 | 15 |
| Total. | 1,001 | 282 | 365 | 46 | 72 | 765 | 15 |
LINKAGE OF SHIFTED, VERMILION, AND BAR.
LINKAGE OF SHIFTED, VERMILION, AND BAR.
In order to determine on which side of vermilion shifted lies, a shifted vermilion (not-bar) female was crossed to a (not-shifted red) bar male. Three factors are involved, of which one, bar, is dominant. The shifted vermilion (not-bar) stock is a triple recessive, and a three-point back-cross was therefore possible. The daughters were bar and the sons were shifted vermilion (the triple recessive). Inbred these gave the results shown in table 46. The smallest classes (double cross-overs) are shifted and vermilion bar, which places shifted to the left of vermilion at approximately 17.8 units from yellow.
To find out where shifted vermilion lies, a shifted vermilion (not-bar) female was bred with a (not-shifted red) bar male. There are three factors involved, one of which, bar, is dominant. The shifted vermilion (not-bar) stock is a triple recessive, allowing for a three-point back-cross. The daughters were bar and the sons were shifted vermilion (the triple recessive). When inbred, these produced the results shown in table 46. The smallest classes (double cross-overs) are shifted and vermilion bar, which places shifted to the left of vermilion at about 17.8 units from yellow.
Table 47.—P1 shifted vermilion ♀ × bar ♂ ♂. F1 bar ♀ × F1 shifted vermillion ♂ ♂.
Table 47.—P1 shifted vermilion female × bar male male. F1 bar female × F1 shifted vermilion male male.
| Reference. | 
| 
| 
| 
| Total. | Cross-over values. | ||||||
| Shifted ver- milion. | Bar. | Shifted bar. | Ver- milion | Shifted ver- milion bar. | Wild- type. | Shifted. | Ver- milion bar. | Shifted ver- milion. | Ver- milion bar. | Shifted bar. | ||
| 65 | 56 | 108 | 15 | 20 | 8 | 33 | 1 | 1 | 242 | 15 | 18 | 31 |
The stock of shifted has been thrown away, since too great difficulty was encountered in maintaining it, because, apparently, of sterility in the females.
The shifted stock has been discarded because it was too difficult to maintain, likely due to infertility in the females.
LETHALS SA AND SB.
LETHALS A and B.
The first lethal found by Miss Rawls was in a stock that had been bred for about 3 years. While there was no a priori reason that could be given to support the view that lethal mutations would occur more frequently among flies inbred in confinement, nevertheless a hundred females from each of several newly caught and from each of several confined stocks were examined for lethals (Stark, 1915). No lethals were found among the wild stocks, but 4 were found among the confined stocks. Whether this difference is significant is perhaps open to question. The first lethal was found in January 1913, in a stock that had been caught at Falmouth, Massachusetts, in 1911, and had been inbred for 18 months, i.e., for about 50 generations. This lethal, lethal sa, was recessive and behaved like the former lethals, being transmitted by half the females and causing the death of half the sons. The position of this lethal to the X chromosome was found as follows, by means of the cross-over value white lethal sa. Lethal-bearing females were mated to white males and the lethal-bearing daughters were again mated to white males. The white sons (894) were non-cross-overs and the red sons (256) were cross-overs. The percentage of crossing-over {65}is 22.2. A correction of 0.4 unit should be added for double crossing-over, indicating that the locus is 22.6 units from white, or at 23.7.
The first lethal discovered by Miss Rawls was in a stock that had been bred for about 3 years. While there was no a priori reason to believe that lethal mutations would happen more often among flies inbred in confinement, a hundred females from several newly caught stocks and from several confined stocks were examined for lethals (Stark, 1915). No lethals were found among the wild stocks, but 4 were found among the confined stocks. Whether this difference is significant is probably up for debate. The first lethal was identified in January 1913, in a stock that had been caught in Falmouth, Massachusetts, in 1911, and had been inbred for 18 months, i.e., for about 50 generations. This lethal, labeled lethal sa, was recessive and acted like the previous lethals, being transmitted by half of the females and causing the death of half the sons. The position of this lethal relative to the X chromosome was determined using the crossover value of white lethal sa. Lethal-bearing females were mated to white males, and the lethal-bearing daughters were then mated to white males again. The white sons (894) were non-crossovers and the red sons (256) were crossovers. The percentage of crossing-over {65}is 22.2. A correction of 0.4 units should be added for double crossing-over, indicating that the locus is 22.6 units from white, or at 23.7.
When the work on lethal sa had been continued for 3 months, the second lethal, lethal sb, was found (April 1913) to be present in a female which was already heterozygous for lethal sa. It is probable that this second lethal arose as a mutation in the father, and that a sperm whose X carried lethal sb fertilized an egg whose X carried lethal sa. As in the cases of lethals 1 and 1a and lethals 3 and 3a, this lethal, lethal sb, was discovered from the fact that only a very few sons were produced, there being 82 daughters and only 3 sons. If, as in the other cases, the number of daughters is taken as the number of non-cross-overs and twice the number of sons as the cross-overs, it is found that the two lethals are about 7 units apart. Since the two lethals were in different X chromosomes, all the daughters should receive one or the other lethal, except in those few cases in which crossing over had taken place. Of the daughters 19 were tested and every one was found to carry a lethal. Again, if the cross-over values of the lethals with some other character, such as white eyes, be found and plotted, the curve should show two modes corresponding to the two lethals. This test was applied, but the curve failed to show two modes clearly,[7] the two lethals being too close together to be differentiated by the small number of determinations that were made. It seems probable that lethal sa and lethal sb are about 5 units apart.
When research on lethal sa had continued for 3 months, a second lethal, lethal sb, was discovered (April 1913) in a female that was already heterozygous for lethal sa. It’s likely that this second lethal resulted from a mutation in the father, and that a sperm with an X chromosome carrying lethal sb fertilized an egg with an X chromosome carrying lethal sa. Just like with lethals 1 and 1a and lethals 3 and 3a, lethal sb was identified because only a very small number of sons were produced, with 82 daughters and only 3 sons. If we consider the number of daughters as the number of non-cross-overs and double the number of sons as the cross-overs, it turns out that the two lethals are about 7 units apart. Since the two lethals were on different X chromosomes, all the daughters should inherit one lethal or the other, except in the rare cases where crossing over occurred. Of the daughters, 19 were tested, and all were found to carry a lethal. Additionally, if the cross-over values of the lethals with another character, like white eyes, are found and plotted, the curve should display two modes corresponding to the two lethals. This test was conducted, but the curve didn't clearly show two modes, as the two lethals were too close together to be distinguished by the small number of determinations made. It seems likely that lethal sa and lethal sb are about 5 units apart.
The position of lethal sb was accurately found by continuing the determinations with a white lethal cross-over. A white female was found which had only one of the two lethals and the linkage of this lethal with eosin and miniature was found as follows: A female carrying white and lethal in one chromosome and no mutant factor in the homologous chromosome was bred to an eosin miniature male. The white eosin daughters carried lethal, and their sons show the amount of crossing-over between white and lethal (15.6), between lethal and miniature (19.9), and between white and miniature (32.9). The data on which these calculations are based are given in table 48.
The lethal sb position was accurately identified by continuing the observations with a white lethal crossover. A white female was found that had only one of the two lethal genes, and the connection of this lethal with eosin and miniature was determined as follows: A female with white and lethal on one chromosome and no mutant factor on the homologous chromosome was bred with an eosin miniature male. The white eosin daughters carried the lethal gene, and their sons showed the amount of crossing-over between white and lethal (15.6), between lethal and miniature (19.9), and between white and miniature (32.9). The data used for these calculations are provided in table 48.
Table 48.—Data on the linkage of white, lethal sb, and miniature, from Stark, 1915.
Table 48.—Data on the connection of white, lethal sb, and miniature, from Stark, 1915.

| 
| 
| 
| Total. | Cross-over values. | ||
| Eosin miniature. | White miniature. | Eosin. | White. | White lethal sb. | Lethal sb. miniature. | White miniature. | |
| 2,421 | 524 | 685 | 48 | 3,678 | 15.6 | 19.9 | 32.9 |
The locus of this lethal is at 16.7; the locus of lethal sa was found to be at 23.7, so that the lethal at 16.7 is evidently the second lethal or lethal sb whose advent gave rise to the high sex-ratio. This interpretation is in accord with the curve which Miss Stark published, for although the mode which corresponds to lethal sa is weak, the mode at 15-16 is well marked.
The location of this lethal is at 16.7; the location of lethal sa was found to be at 23.7, so the lethal at 16.7 is clearly the second lethal or lethal sb, which caused the high sex ratio. This interpretation aligns with the graph that Miss Stark published, because while the mode related to lethal sa is weak, the mode at 15-16 is clearly defined.
The two other lethals, lethals sc and sd, which came up in the course of these experiments by Miss Stark, are treated in other sections of this paper.
The two other lethals, lethals sc and sd, which arose during these experiments by Miss Stark, are discussed in other sections of this paper.
BAR.
Bar.
(Plate II, figures 12 and 13.)
(__A_TAG_PLACEHOLDER_0__, figures 12 and 13.)
The dominant sex-linked mutant called bar-eye (formerly called barred) appeared in February 1913 in an experiment involving rudimentary and long-winged flies (Tice, 1914). A female that is heterozygous for bar has an eye that is intermediate between the rounded eye of the wild fly and the narrow band of the bar stock. This heterozygous bar female is always readily distinguishable from the normal, but can not always be separated from the pure bar. Bar is therefore nearly always used as a dominant and back-crosses are made with normal males.
The dominant sex-linked mutant known as bar-eye (formerly called barred) appeared in February 1913 during an experiment with basic and long-winged flies (Tice, 1914). A female that is heterozygous for bar has an eye that looks like a cross between the rounded eye of the wild fly and the narrow band of the bar stock. This heterozygous bar female is always easy to tell apart from the normal, but she can't always be distinguished from the pure bar. As a result, bar is almost always used as a dominant trait, and back-crosses are done with normal males.
Bar is the most useful sex-linked character so far discovered, on account of its dominance, the certainty of its classification, and its position near the right end of the X chromosome. The locus of bar at 57 was determined on the basis of the data of table 65.
Bar is the most valuable sex-linked trait that has been found so far, due to its dominance, the clarity of its classification, and its location near the right end of the X chromosome. The position of bar at 57 was established based on the data in table 65.
NOTCH.
NOTCH.
A sex-linked dominant factor that brings about a notch at the ends of the wings appeared in March 1913, and has been described and figured by Dexter (1914, p. 753, and fig. 13, p. 730). The factor acts as a lethal for the male. Consequently a female heterozygous for notch bred to a wild male gives a 2:1 sex-ratio; half of her daughters are notch and half normal; the sons are only normal. The actual figures obtained by Dexter were 235 notch females, 270 normal females, and 235 normal males.
A sex-linked dominant trait that causes a notch at the tips of the wings was observed in March 1913, and has been described and illustrated by Dexter (1914, p. 753, and fig. 13, p. 730). This trait is lethal for males. As a result, when a female heterozygous for the notch is bred with a wild male, the sex ratio is 2:1; half of her daughters are notch and half are normal, while all the sons are normal. The actual numbers recorded by Dexter were 235 notch females, 270 normal females, and 235 normal males.
DEPRESSED.
Feeling down.
Several mutations have appeared in which the wings are not flat. Of these the first that appeared was curved (second chromosome), in which the wings are curved downward throughout their length, but are elevated and held out sidewise from the body; the texture is thinner than normal. The second of these wing mutants to appear was jaunty (second chromosome), in which the wings turn up sharply at the tip; they lie in the normal position. The third mutant, arc (second chromosome), has, as its name implies, its wings curved like the arc of a circle. The fourth mutant, bow (first chromosome, fig. c), is like arc, but the amount of curvature is slightly less. The fifth mutant, depressed (first chromosome, fig. g), has the tip of its wings turned down instead of up, as in jaunty, but, as in jaunty, the wing is straight, except near the tip, where it bends suddenly. These stocks have been kept separate since their origin, and flies from them have seldom been crossed to each other, because in the succeeding generations it would be almost impossible to make a satisfactory classification of the various types. But that they are genetically different mutations is at once shown on crossing any two, when wild-type offspring are produced. For instance, bow and arc are the two most nearly alike. Mated together (bow ♂ by arc ♀), they give in F1 straight-winged flies which inbred give in F2 9 straight to 7 not-straight (i.e., bow, arc, and bow arc together).
Several mutations have emerged in which the wings are not flat. The first to appear was the curved mutation (second chromosome), where the wings curve downward along their length but are elevated and extended sideways from the body; the texture is thinner than normal. The second mutant to emerge was the jaunty mutation (second chromosome), characterized by wings that sharply turn up at the tip while remaining in the normal position. The third mutant, arc (second chromosome), has wings that curve like the arc of a circle, as the name suggests. The fourth mutant, bow (first chromosome, fig. c), is similar to arc, but the curvature is slightly less pronounced. The fifth mutant, depressed (first chromosome, fig. g), features wings with the tips turned down instead of up, like in jaunty, but, like jaunty, the wings are straight except near the tip, where they bend suddenly. These stocks have been kept separate since their emergence, and flies from them have rarely been crossed with each other because it would be nearly impossible to make a satisfactory classification of the various types in the following generations. However, their genetic differences become evident when any two are crossed, producing wild-type offspring. For example, bow and arc are the two most similar. When mated together (bow ♂ by arc ♀), they produce straight-winged flies in F1, which, when inbred, yield 9 straight to 7 non-straight in F2 (i.e., bow, arc, and bow arc together).
Depressed wings first appeared (April 1913) among the males of a culture of black flies. They were mated to their sisters and from subsequent generations both males and females with depressed wings were obtained which gave a pure stock. This new character proved to be another sex-linked recessive.
Depressed wings first showed up (April 1913) in the male black flies of a culture. They were bred with their sisters, and from the following generations, both male and female flies with depressed wings were produced, creating a pure stock. This new trait turned out to be another sex-linked recessive.
LINKAGE OF DEPRESSED AND BAR.
LINKAGE OF DEPRESSED AND BAR.
Depressed (not-bar) males mated to (not-depressed) bar females gave bar daughters. Two of these were back-crossed singly to depressed males and gave the results shown in table 49. Males and females were not separated, since they should give the same result.
Depressed (not-bar) males paired with (not-depressed) bar females produced bar daughters. Two of these daughters were back-crossed individually to depressed males, resulting in the findings shown in table 49. Males and females were kept together, as they were expected to yield the same results.
Table 49.—P1 depressed ♀ ♀ × bar ♀ ♀. B.C. F1 bar ♀ × depressed ♂ ♂.
Table 49.—P1 depressed female × bar female. B.C. F1 bar female × depressed male.
| Reference. | Non-cross-overs. | Cross-overs. | Total. | Cross- over values. | ||
| Depressed. | Bar. | Depressed bar. | Wild- type. | |||
| 66 I | 48 | 51 | 21 | 41 | 161 | 39 |
| 67 I | 85 | 104 | 44 | 70 | 303 | 38 |
| Total. | 133 | 155 | 65 | 111 | 464 | 38 |
 Fig. G.—Depressed wing.
Fig. G.—Depressed wing.
LINKAGE OF CHERRY, DEPRESSED, AND VERMILION.
LINKAGE OF CHERRY, DEPRESSED, AND VERMILION.
The linkage value 38 (see table 49) indicates that depressed is somewhere near the opposite end of the series of sex-linked factors from bar. The locus could be more accurately determined by finding the linkage relations of depressed with gens at its end of the chromosome. Accordingly, depressed females were crossed to cherry vermilion males. F1 gave wild-type females and depressed males. The daughters bred again to cherry vermilion males gave the results shown in table 50. The data only suffice to show that the locus of depressed is about midway between cherry and vermilion, or at about 15 units from yellow.
The linkage value 38 (see table 49) indicates that depressed is located near the opposite end of the series of sex-linked factors from bar. The exact location could be better determined by examining the linkage relationships of depressed with genes at the end of the chromosome. So, depressed females were bred with cherry vermilion males. The F1 generation produced wild-type females and depressed males. The daughters were then bred again with cherry vermilion males, resulting in the data shown in table 50. The information is only enough to demonstrate that the locus of depressed is approximately halfway between cherry and vermilion, or about 15 units from yellow.
The F1 males in the last experiment did not have their wings as much depressed as is the condition in stock males, and in F2 most of the depressed winged males were of the F1 type, although a few were like those of stock. This result suggests that the stock is a double recessive, i. e., one that contains, in addition to the sex-linked depressed, an autosomal factor that intensifies the effect of the primary sex-linked factor.
The F1 males in the last experiment didn't have their wings as much drooped as the stock males did, and in the F2, most of the males with drooped wings resembled the F1 type, although a few were similar to the stock. This result suggests that the stock is a double recessive, i.e., one that includes, along with the sex-linked drooped trait, an autosomal factor that amplifies the effect of the main sex-linked factor.
Table 50.—P1 depressed ♀ × cherry vermilion ♂ ♂.
Table 50.—P1 depressed female × cherry vermilion male males.
| First generation. | Second generation. | |||||||||||
| Ref- erence. | ♀♀. | 
| 
| 
|  | |||||||
| Wild- type ♀♀. | Depressed ♂♂. | Cherry ver- milion ♂. | De- pressed ♂. | Cherry de- pressed ♂. | Ver- milion ♂. | Cherry ♂. | De- pressed ver- milion ♂. | Cherry de- pressed ver- milion ♂. | Wild- type ♂. | |||
| 21 | 31 | 19 I | 59 | 23 | 24 | 6 | 6 | 5 | 5 | 0 | 0 | |
CLUB.
GROUP.
In May 1913 there were observed in a certain stock some flies which, although mature, did not unfold their wings (text-fig. Ha). This condition was at first found only in males and suspicion was aroused that the character might be sex-linked. When these males were bred to wild females the club-shaped wings reappeared only in the F2 males, but in smaller number than expected for a recessive sex-linked character. The result led to the further suspicion that not all those individuals that are genetically club show club somatically. These points are best illustrated and proven by the following history of the stock:
In May 1913, some flies were observed in a specific stock that, despite being fully developed, did not spread their wings (text-fig. Ha). Initially, this condition was only found in males, raising the suspicion that it might be linked to sex. When these males were bred with wild females, the club-shaped wings only reappeared in the F2 males, but in fewer numbers than expected for a recessive sex-linked trait. This result led to the further suspicion that not all individuals that are genetically club appear club morphologically. These points are best illustrated and proven by the following history of the stock:

Fig. H.—Club wing. a shows the unexpanded wings of club flies; c shows the absence of the two large bristles from the side of the thorax present in the normal condition of the wild, b.
Fig. H.—Club wing. a shows the undeveloped wings of club flies; c shows the lack of the two large bristles on the side of the thorax that are present in the normal condition of the wild, b.
Club females by wild males gave in the F2 generation (mass cultures): wild-type ♀, 1,131; wild-type ♂, 897; club ♀, 57; club ♂, 131.
Club females from wild males resulted in the F2 generation (mass cultures): wild-type ♀, 1,131; wild-type ♂, 897; club ♀, 57; club ♂, 131.
It is noticeable that there were fewer club females than club males, equality being expected, which might appear to indicate that the club condition is more often realized by the male than by the female, but later crosses show that the difference here is not a constant feature of the cross.
It’s clear that there were fewer female members in the club than male members, which one would expect to be equal. This might suggest that being part of the club is more often achieved by males than females, but later crosses show that this difference isn’t a consistent trait of the cross.
Long-winged males from club stock (potential clubs) bred to wild females gave in F2 the following: wild-type ♀, 521; wild-type (and potential club) ♂, 403; club ♂, 82.
Long-winged males from club stock (potential clubs) bred with wild females produced in F2 the following: wild-type ♀, 521; wild-type (and potential club) ♂, 403; club ♂, 82.
Club females by club males of club stock gave in F2: potential club ♀, 126; potential club ♂, 78; club ♀, 95; club ♂, 81. These results are from 8 pairs. The high proportion of club is noticeable.
Club females bred by club males of club stock produced in F2: potential club ♀, 126; potential club ♂, 78; club ♀, 95; club ♂, 81. These results come from 8 pairs. The high percentage of club is striking.
Potential club females and males from pure club stock (i. e., stock derived originally from a pair of club) gave in F2 the following: potential club ♀, 1,049; potential club ♂, 666; club ♀, 450; club ♂, 453.
Potential club females and males from pure club breeding (i.e., breeding that originally comes from a pair of club) produced in F2 the following: potential club ♀, 1,049; potential club ♂, 666; club ♀, 450; club ♂, 453.
GENOTYPIC CLUB.
GENOTYPE CLUB.
Accurate work with the club character was made possible by the discovery of a character that is a constant index of the presence of homozygous club. This character is the absence of the two large bristles (text-fig. Hc) that are present on each side of the thorax of the wild fly as shown in figure Hb. All club flies are now classified by this character and no attention is paid to whether the wings remain as pads or become expanded.
Accurate work with the club character was made possible by the discovery of a trait that consistently indicates the presence of homozygous club. This trait is the absence of the two large bristles (text-fig. Hc) found on each side of the thorax of the wild fly, as shown in figure Hb. All club flies are now categorized by this trait, and no attention is given to whether the wings remain as pads or expand.
LINKAGE OF CLUB AND VERMILION.
LINK BETWEEN CLUB AND VERMILION.
The linkage of club and vermilion is shown by the cultures listed in table 51, which were obtained as controls in working with lethal III. The cross-over value is shown in the male classes by the cross-over fraction 276/1463 or 19 per cent.
The connection between club and vermilion is demonstrated by the cultures listed in table 51, which were acquired as controls when working with lethal III. The crossover value is indicated in the male classes by the crossover fraction 276/1463, or 19 percent.
LINKAGE OF YELLOW, CLUB, AND VERMILION.
LINKAGE OF YELLOW, CLUB, AND VERMILION.
The data just given in table 51 show that club is 19 units from vermilion, but in order to determine in which direction from vermilion it lies, the crossing-over of club to one other gen must be tested. For this test we used yellow, which lies at the extreme left of the chromosome series. At the same time we included vermilion, so that a three-point experiment was made.
The data presented in table 51 shows that club is 19 units from vermilion, but to figure out which direction it lies from vermilion, the crossing-over of club to another gene needs to be tested. For this test, we used yellow, which is at the far left of the chromosome series. We also included vermilion, so a three-point experiment was conducted.
Females that were (gray) club vermilion were bred to yellow (not-club red) and gave wild-type daughters and club vermilion sons. These inbred gave the results of table 52.
Females that were (gray) club vermilion were bred to yellow (not-club red) and produced wild-type daughters and club vermilion sons. These inbred produced the results in table 52.
The data from the males show that the locus of club is about midway between yellow and vermilion. This conclusion is based on the {71}evidence that yellow and club give 18 per cent of crossing-over, club and vermilion 20 per cent, and yellow and vermilion 35 per cent. The double cross-overs on this view are yellow club (3) and vermilion (3). The females furnish additional data for the linkage of club and vermilion. The value calculated from the female classes alone is 20 units, which is the same value as that given by the males.
The data from the male subjects indicate that the location of the club is roughly halfway between yellow and vermilion. This conclusion is based on the {71}evidence showing that yellow and club result in 18 percent crossing-over, club and vermilion 20 percent, and yellow and vermilion 35 percent. The double crossovers in this analysis include yellow club (3) and vermilion (3). The female subjects provide additional information regarding the connection between club and vermilion. The value calculated solely from the female classes is 20 units, which matches the value reported by the males.
Table 51.—P1 club ♀ ♀ × vermilion ♂ ♂. F1 wild-type ♀ × F1 club ♂.
Table 51.—P1 club female female × vermilion male male. F1 wild-type female × F1 club male.
| Reference. | Females. | Non-cross-over ♂ ♂. | Cross-over ♂ ♂. | Total ♂ ♂. | Cross- over values. | ||
| Club. | Ver- milion. | Club Vermilion. | Wild- type. | ||||
| 137 | 75 | 17 | 39 | 6 | 11 | 73 | 23 |
| 138 | 64 | 24 | 32 | 6 | 8 | 70 | 20 |
| 139 | 56 | 10 | 31 | 4 | 3 | 48 | 15 |
| 140 | 74 | 13 | 39 | 3 | 5 | 60 | 13 |
| 144 | 97 | 30 | 40 | 10 | 13 | 93 | 25 |
| 145 | 63 | 15 | 29 | 4 | 6 | 54 | 19 |
| 146 | 126 | 44 | 46 | 9 | 9 | 108 | 15 |
| 106 | 92 | 33 | 34 | 6 | 10 | 83 | 19 |
| 107 | 55 | 31 | 25 | 7 | 3 | 66 | 15 |
| 108 | 86 | 29 | 32 | 7 | 10 | 78 | 22 |
| 109 | 103 | 25 | 36 | 4 | 9 | 74 | 18 |
| 83 | 30 | 34 | 6 | 9 | 79 | 19 | |
| 77 | 18 | 26 | 7 | 8 | 59 | 25 | |
| 67 | 20 | 21 | 6 | 7 | 54 | 24 | |
| 126 | 32 | 60 | 15 | 13 | 120 | 23 | |
| 63 | 21 | 28 | 7 | 10 | 66 | 26 | |
| 114 | 45 | 71 | 9 | 7 | 132 | 12 | |
| 46 | 18 | 18 | 3 | 3 | 42 | 14 | |
| 111 | 35 | 56 | 6 | 7 | 104 | 13 | |
| Total. | 1,578 | 490 | 697 | 125 | 151 | 1,463 | 19 |
Table 52.—P1 club vermilion ♀ ♀ × yellow ♂ ♂. F1 wild-type ♀ ♀ × F1 club vermilion ♂ ♂.
Table 52.—P1 club vermilion female female × yellow male male. F1 wild-type female female × F1 club vermilion male male.
| Reference. | F2 females. | F2 males. | ||||||||||
| Non-cross- overs. | Cross-overs. | 
| 
| 
|  | |||||||
| Club ver- milion. | Wild- type. | Club. | Ver- milion. | Yellow. | Club ver- milion. | Yellow club ver- milion. | Wild- type. | Yellow ver- milion. | Club. | Yellow Club. | Ver- milion. | |
| 99 | 44 | 52 | 13 | 7 | 35 | 27 | 2 | 9 | 8 | 11 | 0 | 1 |
| 100 | 38 | 58 | 6 | 12 | 43 | 23 | 1 | 15 | 11 | 14 | 0 | 0 |
| 101 | 30 | 32 | 6 | 12 | 19 | 24 | 6 | 5 | 10 | 3 | 1 | 0 |
| 102 | 44 | 55 | 20 | 13 | 48 | 38 | 12 | 14 | 8 | 15 | 1 | 1 |
| 103 | ... | ... | ... | ... | 43 | 32 | 7 | 16 | 13 | 7 | 1 | 1 |
| Total. | 156 | 197 | 45 | 44 | 188 | 144 | 28 | 59 | 50 | 50 | 3 | 3 |
LINKAGE OF CHERRY, CLUB, AND VERMILION.
LINKAGE OF CHERRY, CLUB, AND VERMILION.
The need for a readily workable character whose gen should lie in the long space between cherry and vermilion has long been felt. Cherry and vermilion are so far apart that there must be considerable double crossing-over between them. But with no favorably placed character which is at the same time viable and clearly and rapidly distinguishable, we were unable to find the exact amount of double crossing-over, and hence could not make a proper correction in plotting the chromosome. Club occupies just this favorable position nearly midway between cherry and vermilion. The distances from cherry to club and from club to vermilion are short enough so that no error would be introduced if we ignored the small amount of double crossing-over within each of these distances.
The need for a workable character that fits in the long space between cherry and vermilion has been recognized for a while. Cherry and vermilion are so distinct that a significant amount of double crossing-over must occur between them. However, without a suitable character that is viable and easily distinguishable, we couldn't determine the exact level of double crossing-over, which hindered our ability to accurately plot the chromosome. Club is positioned just right, roughly midway between cherry and vermilion. The distances from cherry to club and from club to vermilion are short enough that we could ignore the small amount of double crossing-over that occurs within these distances without introducing any error.
It thus becomes important to know very exactly the cross-over values for cherry club and club vermilion. The experiment has the form of the yellow club vermilion cross of table 52, except that cherry is used instead of yellow. Cherry is better than yellow because it is slightly nearer club than is yellow and because the bristles of yellow flies are very inconspicuous. In yellow flies the bristles on the side of the thorax are yellowish brown against a yellow background, while in gray-bodied flies the bristles are very black against a light yellowish-gray background.
It’s important to know the exact crossover values for cherry club and club vermilion. The experiment follows the yellow club vermilion cross shown in table 52, but uses cherry instead of yellow. Cherry is better than yellow because it’s closer to club, and the bristles on yellow flies are hard to see. In yellow flies, the bristles on the sides of the thorax are yellowish brown against a yellow background, while in gray-bodied flies, the bristles are very black against a light yellowish-gray background.
For the time being we are able to present only incomplete results upon this cross. In the first experiment cherry females were crossed to club vermilion males and the wild-type daughters were back-crossed to cherry club vermilion, which triple recessive had been secured for this purpose. Table 53 gives the results.
For now, we can only share partial results on this cross. In the first experiment, cherry females were crossed with club vermilion males, and the wild-type daughters were back-crossed to cherry club vermilion, which was secured as triple recessive for this purpose. Table 53 shows the results.
Table 53.—P1 cherry ♀ ♀ × club vermilion ♂ ♂. B. C. F1 wild-type ♀ × cherry club vermilion ♂ ♂.
Table 53.—P1 cherry female × club vermilion male. B. C. F1 wild-type female × cherry club vermilion male.
| Refer- ence. | 
| 
| 
| 
| Total. | Cross-over values. | ||||||
| Cherry. | Club ver- milion. | Cherry club ver- milion. | Wild- type. | Cherry ver- milion. | Club. | Cherry club. | Ver- milion. | Cherry club. | Club ver- milion. | Cherry ver- milion. | ||
| 163 | 68 | 68 | 4 | 10 | 21 | 13 | 1 | 0 | 185 | 8 | 19 | 26 |
| 164 | 99 | 67 | 13 | 21 | 21 | 12 | 1 | 0 | 234 | 15 | 15 | 29 |
| 165 | 23 | 37 | 9 | 7 | 15 | 2 | 0 | 2 | 95 | 19 | 25 | 35 |
| 166 | 107 | 86 | 14 | 28 | 31 | 43 | 3 | 3 | 315 | 15 | 25 | 37 |
| 167 | 42 | 49 | 7 | 11 | 12 | 11 | 2 | 2 | 136 | 16 | 20 | 30 |
| 168 | 40 | 30 | 6 | 15 | 16 | 8 | 0 | 0 | 115 | 18 | 21 | 39 |
| Total. | 379 | 337 | 53 | 92 | 116 | 89 | 7 | 7 | 1,080 | 15 | 20 | 32 |
A complementary experiment was made by crossing cherry club vermilion females to wild males and inbreeding the F1 in pairs. Table 54 gives the results of this cross.
A complementary experiment was conducted by crossing cherry club vermilion females with wild males and inbreeding the F1 in pairs. Table 54 shows the results of this cross.
Table 54.—P1 cherry club vermilion ♂ ♂. ♀ ♀ × wild ♂ ♂. F1 wild-type ♀ × F1 cherry club vermilion ♂ ♂.
Table 54.—P1 cherry club vermilion male male. female female × wild male male. F1 wild-type female × F1 cherry club vermilion male male.
| Refer- ence. | 
| 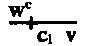
| 
| 
| Total. | Cross-over values. | ||||||
| Cherry club ver- milion. | Wild- type. | Cherry. | Club ver- milion. | Cherry club. | Ver- milion. | Cherry ver- milion. | Club. | Cherry club. | Club ver- milion. | Cherry ver- milion. | ||
| 188 | 60 | 76 | 12 | 8 | 12 | 29 | 2 | 1 | 200 | 11 | 22 | 30 |
| 189 | 228 | 314 | 48 | 44 | 50 | 60 | 1 | 8 | 753 | 13 | 16 | 27 |
| 197 | 68 | 81 | 23 | 13 | 9 | 22 | 2 | 0 | 218 | 17 | 15 | 31 |
| Total. | 356 | 471 | 83 | 65 | 71 | 111 | 5 | 9 | 1,171 | 14 | 17 | 28 |
The combined data of tables 53 and 54 give 14.2 as the value for cherry club. All the data thus far presented upon club vermilion (886 cross-overs in a total of 4,681), give 19.2 as the value for club vermilion. The locus of club on the basis of the total data available is at 14.6.
The combined data from tables 53 and 54 shows a value of 14.2 for cherry club. All the data presented so far for club vermilion (886 cross-overs out of a total of 4,681) gives a value of 19.2 for club vermilion. Based on all the available data, the locus of club is at 14.6.
GREEN.
GREEN.
In May 1913 there appeared in a culture of flies with gray body-color a few males with a greenish-black tinge to the body and legs. The trident pattern on the thorax, which is almost invisible in many wild flies, was here quite marked. A green male was mated to wild females and gave in F2 a close approach to a 2:1:1 ratio. The green reappeared only in the F2 males, but the separation of green from gray was not as easy or complete as desirable. From subsequent generations a pure stock of green was made. A green female by wild male gave 138 wild-type females and 127 males which were greenish. This green color varies somewhat in depth, so that some of these F1 males could not have been separated with certainty from a mixed culture of green and gray males.
In May 1913, a culture of flies with gray bodies had a few males that had a greenish-black tint on their bodies and legs. The trident pattern on the thorax, which is almost invisible in many wild flies, was quite prominent here. A green male was paired with wild females and produced an F2 generation that closely resembled a 2:1:1 ratio. The green coloration came up again only in the F2 males, but distinguishing between green and gray wasn't as straightforward or effective as hoped. From later generations, a pure green stock was established. A green female mated with a wild male produced 138 wild-type females and 127 males that were greenish. This green color varies in intensity, so some of these F1 males might not have been reliably distinguished from a mixed group of green and gray males.
The results of these two experiments show that green is a sex-linked melanistic character like sable, but the somatic difference produced is much less than in the case of sable, so that the new mutation, although genetically definite, is of little practical value. We have found several eye-colors which differed from the red color of the wild fly by very small differences. With some of these we have worked successfully by using another eye-color as a developer. For example, the double recessive vermilion "clear" is far more easily distinguished from vermilion than is clear from red. But it is no small task to make up the stocks {74}necessary for such a special study. In the case of green we might perhaps have employed a similar method, performing all experiments with a common difference from the gray in all flies used.
The results of these two experiments show that green is a sex-linked melanistic trait like sable, but the somatic difference it produces is much less significant than in the case of sable, meaning that the new mutation, although genetically distinct, is of little practical value. We have found several eye colors that differ from the red color of the wild fly by only small differences. With some of these, we have successfully worked by using another eye color as a developer. For example, the double recessive vermilion "clear" is much easier to distinguish from vermilion than clear is from red. However, creating the necessary stocks for such a specific study is no small task. In the case of green, we might have been able to use a similar method, conducting all experiments with a common difference from the gray in all flies used.
CHROME.
Chrome.
In a stock of forked fused there appeared, September 15, 1913, three males of a brownish-yellow body-color. They were uniform in color, without any of the abdominal banding so striking in other body-colors. Even the tip of the abdomen lacked the heavy pigmentation which is a marked secondary sexual character of the male. About 20 or more of these males appeared in the same culture. This appearance of many males showing a mutant character and the non-appearance of corresponding females is usual for sex-linked characters. In such cases females appear in the next generation, as they did in this case when the chrome males were mated to their sisters in mass cultures. Since both females and males of chrome were on hand, it should have been an easy matter to continue the stock, but many matings failed, and it was necessary to resort to breeding in heterozygous form. The chrome, however, gradually disappeared from the stock. Such a difficult sex-linked mutation as this could be successfully handled (like a lethal) if it could be mated to a double recessive whose members lie one on each side of the mutant, but in the case of chrome this was not attempted soon enough to save the stock.
On September 15, 1913, a stock of forked fused showed three males with a brownish-yellow body color. They all looked the same, lacking the abdominal banding that is so noticeable in other colors. Even the tip of the abdomen was missing the heavy pigmentation that is a typical secondary sexual feature of males. Around 20 or more of these males appeared in the same culture. It’s common for many males with a mutant trait to appear while corresponding females do not, as this is characteristic of sex-linked traits. Generally, females appear in the next generation, which happened here when the chrome males were mated with their sisters in mass cultures. Since both chrome males and females were available, it should have been straightforward to continue the stock, but many matings didn’t succeed, necessitating breeding in a heterozygous form. However, the chrome gradually faded from the stock. Handling such a challenging sex-linked mutation could have been managed (like a lethal) if it had been mated to a double recessive that was on each side of the mutant, but this approach was not attempted soon enough to save the stock.
LETHAL 3.
LETHAL 3.
In the repetition of a cross between a white miniature male and a vermilion pink male (December 1913), the F2 ratios among the males were seen to be very much distorted because of the partial absence of certain classes (Morgan 1914c). While it was suspected that the disturbance was due to a lethal, the data were useless for determining the position of such a lethal, from the fact that more than one mother had been used in each culture. From an F2 culture that gave practically a 2:1 sex-ratio, vermilion females were bred to club males. Several such females gave sex-ratios. Their daughters were again mated to vermilion males. Half of these daughters gave high female sex-ratios and showed the linkage relations given in table 55.
In a repeat experiment involving a cross between a white miniature male and a vermilion pink male (December 1913), the F2 ratios among the males were significantly distorted due to the partial absence of certain classes (Morgan 1914c). Although it was suspected that the disruption was caused by a lethal factor, the data were inadequate for pinpointing the location of such a lethal because multiple mothers had been used in each culture. From an F2 culture that resulted in almost a 2:1 sex ratio, vermilion females were bred with club males. Several of these females produced sex ratios. Their daughters were again bred with vermilion males. Half of these daughters yielded high female sex ratios and displayed the linkage relations outlined in table 55.
Table 55.—Linkage data on club, lethal 3, and vermilion, from Morgan, 1914c.
Table 55.—Linkage data on club, lethal 3, and vermilion, from Morgan, 1914c.
| Females. | Males. | |||

| 
| 
|  | |
| Club. | Wild-type. | Club vermilion. | Vermilion. | |
| 588 | 182 | 28 | 11 | 1 |
Lethal 3 proved to lie between club and vermilion, 13 units from club and 5 from vermilion. The same locus was indicated by the data from the cross of vermilion lethal-bearing females by eosin miniature males. The complete data bearing on the position of lethal 3 is summarized in table 56. On the basis of this data lethal 3 is located at 26.5.
Lethal 3 is positioned between club and vermilion, 13 units away from club and 5 units away from vermilion. The same location was identified by the data from the cross of vermilion lethal-carrying females and eosin miniature males. The complete data regarding the position of lethal 3 is summarized in table 56. Based on this data, lethal 3 is located at 26.5.
Table 56.—Summary of linkage data on lethal 3, from Morgan, 1914c.
Table 56.—Summary of linkage data on lethal 3, from Morgan, 1914c.
| Gens. | Total. | Cross- overs. | Cross-over values. |
| Eosin lethal 3 | 1,327 | 268 | 20.2 |
| Eosin vermilion | 1,327 | 357 | 27.0 |
| Eosin miniature | 3,374 | 967 | 29.0 |
| Club lethal 3 | 222 | 29 | 13.0 |
| Club vermilion | 877 | 161 | 18.4 |
| Lethal 3 vermilion | 1,549 | 105 | 6.8 |
| Lethal 3 miniature | 1,481 | 138 | 9.3 |
| Vermilion miniature | 1,327 | 31 | 2.3 |
LETHAL 3a.
LETHAL 3a.
In January 1914 a vermilion female from a lethal 3 culture when bred to a vermilion male gave 71 daughters and only 3 sons; 34 of these daughters were tested, and every one of them gave a 2:1 sex-ratio. The explanation advanced (Morgan 1914c) was that the mother of the high ratio was heterozygous for lethal 3, and also for another lethal that had arisen by mutation in the X chromosome brought in by the sperm. On this interpretation the few males that survived were those that had arisen through crossing-over. The rarity of the sons shows that the two lethals were in loci near together, although here of course in different chromosomes, except when one of them crossed over to the other. As explained in the section on lethal 1 and 1a the distance between the two lethals can be found by taking twice the number of the surviving males (2+3) as the cross-overs and the number of the females as the non-cross-overs. But the 34 daughters tested were also non-cross-overs, since none of them failed to carry a lethal. The fractions (6+0)/(71+34) = 6/105 give 5.7 as the distance between the lethals in question. In the case of lethals 3 and 3a another test was applied which showed graphically that two lethals were present. Each of the daughters tested showed, by the classes of her sons, the amount of crossing-over between white and that lethal of the two that she carried. These cross-over values were plotted and gave a bimodal curve with modes 7 units apart. It had already been shown that the locus of one of the two lethals was at 26.5, and since the higher of the two modes was at about 23, it corresponds to lethal 3. The data and the curve show that the lethals 3 and 3a are about 7 units apart, i. e., lethal 3a lies at about 19.5. {76}
In January 1914, a red female from a lethal 3 culture bred to a red male produced 71 daughters and only 3 sons. Of these daughters, 34 were tested, and they all showed a 2:1 sex ratio. The explanation given (Morgan 1914c) was that the mother with the high ratio was heterozygous for lethal 3 and also for another lethal that had mutated on the X chromosome from the sperm. According to this interpretation, the few males that survived were those that formed through crossing-over. The rarity of the sons indicates that the two lethals were located near each other, though on different chromosomes, unless one crossed over to the other. As explained in the section on lethal 1 and 1a, the distance between the two lethals can be determined by taking twice the number of surviving males (2+3) as the cross-overs and the number of females as the non-cross-overs. However, the 34 tested daughters were also non-cross-overs since none of them failed to carry a lethal. The fractions (6+0)/(71+34) = 6/105 yield a distance of 5.7 between the lethals in question. For lethals 3 and 3a, another test was applied which visually confirmed that two lethals were present. Each tested daughter demonstrated, through the classes of her sons, the degree of crossing-over between white and the lethal she carried. These cross-over values were plotted, resulting in a bimodal curve with modes 7 units apart. It had previously been established that the locus of one of the lethals was at 26.5, and since the higher of the two modes was around 23, it corresponds to lethal 3. The data and the curve indicate that lethals 3 and 3a are about 7 units apart, i.e., lethal 3a is located at approximately 19.5. {76}
LETHAL 1b.
LETHAL 1b.
A cross between yellow white males and abnormal abdomen females gave (February 1914) regular results in 10 F2 cultures, but three cultures gave 2 ♀ : 1 ♂ sex-ratios (Morgan, 1914b, p. 92). The yellow white class, which was a non-cross-over class in these 10 cultures, had disappeared in the 3 cultures. Subsequent work gave the data summarized in table 57. At the time when the results of table 57 were obtained it did not seem possible that two different lethals could be present in the space of about 1 unit between yellow and white, and this lethal was thought to be a reappearance of lethal 1 (Morgan, 1912b, p. 92). Since then a large number of lethals have arisen, one of them less than 0.1 unit from yellow, and at least one other mutation has taken place between yellow and white, so that the supposition is now rather that the lethal in question was not lethal 1. Indeed, the linkage data show that this lethal, which may be called lethal 1b, lies extraordinarily close to white, for the distance from yellow was 0.8 unit and of white from yellow on the basis of the same data 0.8. There was also a total absence of cross-overs between lethal 1b and white in the total of 846 flies which could have shown such crossing-over. On the basis of this linkage data alone we should be obliged to locate lethal 1b at the point at which white itself is situated, namely, 1.1, but on a priori grounds it seems improbable that a lethal mutation has occurred at the same locus as the factor for white eye-color. Farther evidence against this supposition is that females that have one X chromosome with both yellow and white and the other X chromosome with yellow, lethal, and white are exactly like regular stock yellow white flies. The lethal must have appeared in a chromosome which was already carrying white and yet did not affect the character of the white. We prefer, therefore, to locate lethal 1b at 1.1-.
A cross between yellow white males and females with abnormal abdomens produced consistent results in 10 F2 cultures, but three cultures showed a 2 ♀ : 1 ♂ sex ratio (Morgan, 1914b, p. 92). The yellow white class, which did not undergo crossover in these 10 cultures, was absent in the 3 cultures. Further research provided the data summarized in table 57. When the results of table 57 were obtained, it didn't seem possible for two different lethals to be present within about 1 unit between yellow and white, and this lethal was thought to be a reappearance of lethal 1 (Morgan, 1912b, p. 92). Since then, many lethals have emerged, one of them located less than 0.1 unit from yellow, and there has been at least one other mutation between yellow and white, leading us to believe that the lethal in question was not lethal 1. Indeed, the linkage data indicate that this lethal, which we can call lethal 1b, is extremely close to white, with a distance of 0.8 unit from yellow, and the distance of white from yellow being the same based on the same data, 0.8. There was also a complete absence of cross-overs between lethal 1b and white among the total of 846 flies that could have displayed such crossing-over. Based solely on this linkage data, we would have to place lethal 1b at the same point as white, which is 1.1, but based on a priori reasoning, it seems unlikely that a lethal mutation has occurred at the same location as the factor for white eye color. Further evidence against this assumption is that females with one X chromosome containing both yellow and white and the other X chromosome with yellow, lethal, and white are identical to regular stock yellow white flies. Therefore, the lethal must have arisen in a chromosome that was already carrying white without affecting the white characteristic. We prefer, therefore, to place lethal 1b at 1.1-.
Table 57.—Summary of all linkage data upon lethal 1b, from Morgan, 1914b.
Table 57.—Summary of all linkage data on lethal 1b, from Morgan, 1914b.
| Gens. | Total. | Cross- overs. | Cross-over values. |
| Yellow lethal 1b | 744 | 6 | 0.81 |
| Yellow white | 2,787 | 23 | 0.82 |
| Lethal 1b white | 846 | 0 | 0.0 |
FACET.
Facet.
Several autosomal mutations had been found in which the facets of the compound eye are disarranged. One that was sex-linked appeared in February 1914. Under the low power of the binocular microscope the facets are seen to be irregular in arrangement, instead of being arranged in a strictly regular pattern. The ommatidia are more nearly circular than hexagonal in outline, and are variable in size, some being considerably larger than normal. The large ones are also darker than {77}the smaller, giving a blotched appearance to the eye. The short hairs between the facets point in all directions instead of radially, as in the normal eye. The irregular reflection breaks up the dark fleck which is characteristic of the normal eye. The shape of the eye differs somewhat from the normal; it is more convex, smaller, and is encircled by a narrow rim destitute of ommatidia.
Several genetic mutations have been discovered that cause the facets of the compound eye to be misaligned. One mutation that was linked to sex appeared in February 1914. Under low power magnification of the binocular microscope, the facets are seen to be arranged irregularly, rather than in a strictly regular pattern. The ommatidia are more circular than hexagonal in shape and vary in size, with some being significantly larger than normal. The larger ones are also darker than the smaller ones, creating a blotchy look to the eye. The short hairs between the facets point in all directions instead of radiating outward, as they do in a normal eye. This irregular reflection disrupts the dark spot that is typical of a normal eye. The overall shape of the eye differs slightly from normal; it is more rounded, smaller, and surrounded by a narrow rim that lacks ommatidia.
Facet arose in a back-cross to test the independence of speck (second chromosome) and maroon (third chromosome). One of the cultures produced, among the first males to hatch, some males which showed the facet disarrangement. None of the females showed this character. The complete output was that typical of a female heterozygous for a recessive sex-linked character: not-facet ♀ ♀ (2), 112; not-facet ♂ ♂ (1), 57; facet ♂ ♂ (1), 51.
Facet emerged from a back-cross to examine the independence of speck (second chromosome) and maroon (third chromosome). One of the cultures produced, among the first males to hatch, some males displaying the facet disarrangement. None of the females exhibited this trait. The overall results were typical of a female heterozygous for a recessive sex-linked trait: not-facet ♀ ♀ (2), 112; not-facet ♂ ♂ (1), 57; facet ♂ ♂ (1), 51.
Of the three characters which were shown by the F2 males, one, facet, is sex-linked, another, speck, is in the second chromosome, and maroon is in the third chromosome. All eight F2 classes are therefore expected to be equal in size, and each pair of characters should show free assortment, that is, 50 per cent. The assortment value for facet speck is 48, for speck maroon 52, and for facet maroon 48, as calculated from the F2 males of table 58.
Of the three traits shown by the F2 males, one, facet, is linked to sex, another, speck, is located on the second chromosome, and maroon is on the third chromosome. Therefore, all eight F2 classes are expected to be of equal size, and each pair of traits should assort independently, meaning 50 percent. The assortment values for facet and speck are 48, for speck and maroon 52, and for facet and maroon 48, based on the F2 males in table 58.
Table 58.—P1 speck maroon ♂ × wild ♀ ♀. B.C. F1 wild-type ♀ × speck maroon ♂.
Table 58.—P1 speck maroon male × wild female female. B.C. F1 wild-type female × speck maroon male.
| Reference. | F2 females. | F2 males. | ||||||||||
| Speck maroon. | Wild- type. | Speck. | Maroon. | Facet. | Speck maroon. | Facet speck maroon. | Wild- type. | Facet maroon. | Speck. | Aspect speck. | Maroon. | |
| 66 | 31 | 30 | 26 | 25 | 14 | 14 | 14 | 10 | 11 | 17 | 12 | 17 |
LINKAGE OF FACET, VERMILION AND SABLE.
LINKAGE OF FACET, VERMILION AND SABLE.
In order to determine the location of facet in the first chromosome, one of the facet males which appeared in culture 66 was crossed out to vermilion sable females. Three of the wild-type daughters were back-crossed to vermilion sable males. The females of the next generation should give data upon the linkage of vermilion and sable, while the males should show the linkage of all three gens, facet, vermilion, and sable. The offspring of these three females are classified in table 59.
In order to find the location of the facet gene on the first chromosome, one of the facet males from culture 66 was crossed with vermilion sable females. Three of the wild-type daughters were back-crossed to vermilion sable males. The females from the next generation should provide information about the linkage of vermilion and sable, while the males should reveal the linkage of all three genes: facet, vermilion, and sable. The offspring of these three females are categorized in table 59.
The cross-over fraction for vermilion sable as calculated from the females is 19/194. The cross-over value corresponding to this fraction is 10 units, which was the value found in the more extensive experiments given in the section on sable.
The cross-over fraction for vermilion sable calculated from the females is 19/194. The cross-over value associated with this fraction is 10 units, which was the value found in the more detailed experiments discussed in the section on sable.
It will be noticed that the results in the males of culture 150 are markedly different from those of the other two pairs. While the sable males are fully represented, their opposite classes, the gray males, are {78}entirely absent. This result is due to a lethal factor, lethal 5, which appeared in this culture for the first time.
It’s clear that the results in the male samples from culture 150 are significantly different from those in the other two pairs. While the sable males are fully present, their counterparts, the gray males, are {78}completely missing. This outcome is caused by a lethal factor, lethal 5, which emerged in this culture for the first time.
The males of the two cultures 149 and 151 give the order of gens as facet, vermilion, sable; that is, facet lies to the left of vermilion and toward yellow. The cross-over values are: facet vermilion 40; vermilion sable 12; facet sable 42. Since yellow and vermilion usually give but 34 per cent of crossing-over, this large value of 40 for facet vermilion shows that facet must lie very near to yellow.
The males from cultures 149 and 151 list the gens in this order: facet, vermilion, sable; meaning, facet is to the left of vermilion and closer to yellow. The crossover values are: facet vermilion 40; vermilion sable 12; facet sable 42. Since yellow and vermilion typically produce only 34 percent crossover, the high value of 40 for facet vermilion indicates that facet must be very close to yellow.
Table 59.—P1 facet ♂ × vermilion sable ♀ ♀. B.C. F1 wild-type ♀ × vermilion sable ♂ ♂.
Table 59.—P1 facet male × vermilion sable female. B.C. F1 wild-type female × vermilion sable male.
| Reference. | F2 females. | F2 males. | ||||||||||
| Non-cross-overs. | Cross-overs. | 
| 
| 
|  | |||||||
| Ver- milion sable. | Wild- type. | Ver- milion. | Sable. | Facet. | Ver- milion sable. | Facet ver- milion sable. | Wild- type. | Facet sable. | Ver- milion. | Facet ver- milion. | Sable. | |
| 149 | 16 | 29 | 3 | 3 | 17 | 10 | 8 | 12 | 2 | .. | 2 | 1 |
| 150 | 13 | 17 | 2 | 2 | .. | 10 | 9 | .. | 1 | .. | .. | .. |
| 151 | 37 | 63 | 7 | 2 | 38 | 23 | 12 | 26 | 2 | 8 | 4 | 1 |
| Total. | 66 | 109 | 12 | 8 | 55 | 43 | 29 | 38 | 5 | 8 | 6 | 2 |
LINKAGE OF EOSIN, FACET, AND VERMILION.
LINKAGE OF EOSIN, FACET, AND VERMILION.
In order to obtain more accurate information on the location of facet, a facet male was mated to an eosin vermilion female. The F1 females were mated singly to wild males and they gave the results shown in table 60. The F2 females were not counted, since they do not furnish any information. The evidence of table 60 places facet at 1.1 units to the right of eosin, or at 2.2.
To get more precise information about the location of the facet, a facet male was paired with an eosin vermilion female. The F1 females were mated individually with wild males, resulting in the findings shown in table 60. The F2 females were not included in the count since they don't provide any relevant information. The data in table 60 indicates that the facet is located 1.1 units to the right of eosin, or at 2.2.
Table 60.—P1 eosin vermilion ♀ × facet ♂. F1 wild-type ♀ × wild ♂.
Table 60.—P1 eosin vermilion ♀ × facet ♂. F1 wild-type ♀ × wild ♂.
| Reference. | 
| 
| 
| 
| Total. | Cross-over values. | ||||||
| Eosin ver- milion. | Facet. | Eosin facet. | Ver- milion. | Eosin. | Facet ver- milion. | Eosin facet ver- milion. | Wild- type. | Eosin facet. | Facet ver- milion. | Eosin ver- milion. | ||
| 512 | 43 | 43 | .. | 1 | 13 | 16 | .. | .. | 116 | .... | .... | .... |
| 513 | 28 | 35 | .. | 2 | 19 | 5 | .. | .. | 89 | .... | .... | .... |
| 514 | 18 | 31 | 1 | .. | 17 | 11 | .. | .. | 78 | .... | .... | .... |
| 515 | 18 | 60 | .. | .. | 20 | 15 | .. | .. | 113 | .... | .... | .... |
| 516 | 10 | 31 | .. | .. | 7 | 12 | .. | .. | 60 | .... | .... | .... |
| 517 | 24 | 34 | .. | .. | 10 | 12 | .. | .. | 80 | .... | .... | .... |
| 518 | 44 | 38 | 1 | 1 | 23 | 22 | .. | 1 | 130 | .... | .... | .... |
| Total. | 185 | 272 | 2 | 4 | 109 | 93 | .. | 1 | 666 | 1.05 | 30.5 | 31.3 |
LETHAL SC.
LETHAL SC.
The third of the lethals which Miss Stark found (Stark, 1915) while she was testing the relative frequency of occurrence of lethals in fresh and inbred wild stocks arose in April 1914 in stock caught in 1910. Females heterozygous for this lethal, lethal sc, were mated to white males and the daughters were back-crossed to white males. Half of the daughters gave lethal sex-ratio, and these gave 1,405 cross-overs in a total of 3,053 males, from which the amount of crossing-over between white and lethal sc has been calculated as 46 per cent.
The third lethal that Miss Stark discovered (Stark, 1915) while she was investigating the relative frequency of lethals in fresh and inbred wild populations appeared in April 1914 in a stock collected in 1910. Females that were heterozygous for this lethal, lethal sc, were paired with white males, and the daughters were back-crossed with white males. Half of the daughters showed a lethal sex ratio, and these produced 1,405 cross-overs out of a total of 3,053 males, which allowed the calculation of the crossing-over between white and lethal sc to be 46 percent.
By reference to table 65 it is seen that white and bar normally give only about 44 per cent of crossing-over in a two-locus experiment; lethal sc then is expected to be situated at least as far to the right as bar. Females heterozygous for lethal sc were therefore crossed to bar males, and their daughters were tested. The lethal-bearing daughters gave 144 cross-overs in a total of 1,734 males, that is, bar and lethal sc gave 8.3 per cent of crossing-over. Lethal sc therefore lies 8.3 units beyond bar or at about 66.5. The cross-over value sable lethal sc was found to be 23.5 (387 cross-overs in a total of 1,641 males) which places the lethal at 43+23.5, or at 66.5. We know from other data that there is enough double crossing-over in the distance which gives an experimental value of 23.5 per cent, so that the true distance is a half unit longer or the locus at 67.0 is indicated by the 1,641 males of the sable lethal experiment. In a distance so short that the experimental value is only 8.3 per cent there is, as far as we have been able to determine, no double crossing-over at all, or at most an amount that is entirely negligible, so that a locus at 57+8.3 or 65.3 is indicated by the 1,734 males of the bar lethal experiment. To get the value indicated by the total data the cases may be weighted, that is, the value 65.3 may be multiplied by 1,734, and 67.0 may be multiplied by 1,641. The sum of these two numbers divided by the sum of 1,734 and 1,641 gives 66.2 as the locus indicated by all the data available. This method has been used in every case where more than one experiment furnishes data upon the location of a factor. In constructing the map given in diagram I rather complex balancings were necessary.
By looking at table 65, we see that white and bar typically result in about 44 percent of crossing-over in a two-locus experiment; thus, lethal sc is expected to be located at least as far to the right as bar. Females that are heterozygous for lethal sc were crossed with bar males, and their daughters were tested. The daughters that carried the lethal gene produced 144 cross-overs out of a total of 1,734 males, meaning that bar and lethal sc resulted in 8.3 percent of crossing-over. Therefore, lethal sc is positioned 8.3 units beyond bar, or at about 66.5. The crossover value for sable lethal sc was found to be 23.5 (with 387 cross-overs out of 1,641 males), which places the lethal at 43 + 23.5, or at 66.5. We know from additional data that there's enough double crossing-over in that distance to give an experimental value of 23.5 percent, so the true distance is half a unit longer, suggesting the locus is at 67.0 as indicated by the 1,641 males in the sable lethal experiment. In such a short distance, where the experimental value is only 8.3 percent, it seems there is no double crossing-over, or at most a negligible amount, indicating a locus at 57 + 8.3 or 65.3, based on the 1,734 males from the bar lethal experiment. To find the value indicated by the total data, the cases can be weighted; the value 65.3 can be multiplied by 1,734, and 67.0 by 1,641. The sum of these two results divided by the total of 1,734 and 1,641 gives 66.2 as the locus indicated by all the available data. This method has been utilized in every case where multiple experiments provide data on the location of a factor. When constructing the map shown in diagram I, a rather complex balancing act was necessary.
LETHAL SD.
LETHAL SD.
The fourth lethal which Miss Stark found (May 1914) in the inbred stocks of Drosophila has not been located by means of linkage experiments. It is interesting in that the males which receive the lethal factor sometimes live long enough to hatch. These males are extremely feeble and never live more than two days. There is, as far as can be seen, no anatomical defect to which their extreme feebleness and early death can be attributed. {80}
The fourth lethal factor that Miss Stark discovered (May 1914) in the inbred stocks of Drosophila hasn't been identified through linkage experiments. It's interesting because the males that inherit this lethal factor sometimes survive long enough to hatch. These males are very weak and never live for more than two days. There doesn't seem to be any anatomical defect that explains their extreme weakness and short lifespan. {80}
FURROWED.
Furrowed.
In studying the effect of hybridization upon the production of mutations in Drosophila, F. N. Duncan found a sex-linked mutation which he called "furrowed eye" (Duncan 1915). The furrowed flies are characterized by a foreshortening of the head, which causes the surface of the eye to be thrown into irregular folds with furrows between. The spines of the scutellum are stumpy, a character which is of importance in classification, since quite often flies occur which have no noticeable disturbance of the eyes.
In studying how hybridization affects the production of mutations in Drosophila, F. N. Duncan discovered a sex-linked mutation he named "furrowed eye" (Duncan 1915). The furrowed flies have a shortened head, which causes the eye's surface to form irregular folds with grooves in between. The spines on the scutellum are short, which is important for classification, as there are often flies that show no noticeable changes in the eyes.
The locus of furrowed was determined to be at 38.0 on the basis of the data given in table 61.
The location of the furrowed region was found to be at 38.0 based on the data provided in table 61.
Table 61.—Data on the linkage of furrowed, from Duncan, 1915.
Table 61.—Data on the linkage of furrowed, from Duncan, 1915.
| Gens. | F2 males. | Total. | Cross-over values. | |||||

| 
| 
| 
| Eosin minia- ture. | Minia- ture fur- rowed. | Eosin fur- rowed. | ||
| Eosin, miniature, furrowed. | 142 | 59 | 4 | 3 | 208 | 29.8 | 30.4 | 30.3 |

| 
| 
| 
| Fur- rowed sable. | Sable forked. | Fur- rowed forked. | ||
| Furrowed, sable, forked. | 166 | 9 | 31 | 3 | 209 | 5.7 | 16.3 | 19.1 |

| 
| 
| 
| Verm- ilion fur- rowed. | Fur- rowed bar. | Verm- ilion fur- rowed. | ||
| Vermilion, fur- rowed, bar. | 188 | 9 | 43 | 0 | 240 | 3.8 | 21.6 | 17.9 |
ADDITIONAL DATA FOR YELLOW, WHITE, VERMILION, AND MINIATURE.
ADDITIONAL DATA FOR YELLOW, WHITE, VERMILION, AND MINIATURE.
Considerable new work has been done by various students upon the linkage of the older mutant characters, namely, yellow, white, vermilion, and miniature. We have summarized these new data, and they give values very close to those already published. We have included in the white miniature data those published by P. W. Whiting (Whiting 1913). {81}
Considerable new research has been conducted by various students on the connections between the older mutant traits, specifically yellow, white, vermilion, and miniature. We’ve summarized this new information, which aligns closely with previous findings. We’ve also included in the white miniature data those published by P. W. Whiting (Whiting 1913). {81}
Table 62.—Data upon the linkage of yellow, white, vermilion, and miniature (contributed by students).
Table 62.—Information on the connection of yellow, white, vermilion, and miniature (provided by students).
| Gens. | Non-cross-overs. | Cross-overs. | Total. | Cross-over values. | ||

|  | |||||
| White miniature. | 6,219[8] | 7,378 | 3,754 | 3,337 | 20,688 | 34.2 |

| 
| |||||
| 1,651 | 1,116 | 671 | 1,047 | 4,485 | 38.3 | |

| 
| |||||
| Yellow miniature. | 761 | 923 | 421 | 653 | 2,758 | 39 |

| 
| |||||
| Vermilion miniature. | 1,685 | 1,460 | 32 | 36 | 3,213 | 2.1 |

| 
| |||||
| Yellow white. | 1,600 | 1,807 | 10 | 7 | 3,424 | 0.5 |

| 
| |||||
| Yellow vermilion. | 509 | 587 | 328 | 284 | 1,708 | 35.8 |

| 
| |||||
| White bar. | 198 | 272 | 168 | 166 | 804 | 42 |

| 
| |||||
| Bifid rudimentary. | 142 | 15 | 12 | 116 | 285 | 45 |
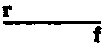
| 
| |||||
| Rudimentary forked. | 73 | 211 | ... | 4 | 288 | 1.4 |
NEW DATA CONTRIBUTED BY A. H. STURTEVANT AND H. J. MULLER.
NEW DATA CONTRIBUTED BY A. H. STURTEVANT AND H. J. MULLER.
Data from several experiments upon sex-linked characters described in this paper have been contributed by Dr. A. H. Sturtevant and Mr. H. J. Muller, and are given in table 63.
Data from several experiments on sex-linked traits described in this paper have been provided by Dr. A. H. Sturtevant and Mr. H. J. Muller, and are included in table 63.
Table 63.—Data contributed by A. H. Sturtevant and H. J. Muller.
Table 63.—Data provided by A. H. Sturtevant and H. J. Muller.
| Gens. | Classes. | Total. | Cross-over values. | ||||||||||||

| 
| 
| 
| Yellow white. | White bifid. | Yellow bifid. | |||||||||
| Yellow white × bifid. | 233 | 254 | 1 | 2 | 10 | 6 | .. | .. | 506 | 0.6 | 3.2 | 3.8 | |||
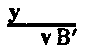
| 
| 
| 
| Yellow ver- milion. | Ver- milion bar. | Yellow bar. | |||||||||
| Yellow × vermilion bar. | 99 | 101 | 60 | 55 | 49 | 48 | 9 | 14 | 435 | 32 | 28 | 49 | |||

| 
| 
| 
| White bifid. | Bifid forked. | White forked. | |||||||||
| White bifid × forked. | 84 | 77 | 9 | 6 | 65 | 59 | 1 | 5 | 306 | 7 | 42 | 45 | |||

| 
| 
| 
| Ver- milion minia- ture. | Minia- ture sable. | Ver- milion sable. | |||||||||
| Vermilion miniature × sable. | 152 | 111 | 4 | 2 | 5 | 12 | .. | .. | 286 | 2.1 | 6 | 8.1 | |||

| 
| 
| 
| Sable rudi- men- tary. | Rudi- men- tary forked. | Sable forked. | |||||||||
| Sable rudimentary × forked. | 143 | 195 | 26 | 27 | 4 | 3 | .. | .. | 398 | 13.3 | 1.8 | 15 | |||
| White Bifid × Basic. | |||||||||||||||
| F2 females. | F2 males. | Total. | Cross-over values. | ||||||||||||

| 
| 
| 
| 
| 
| White bifid. | Bifid rudi- men- tary. | White rudi- men- tary. | |||||||
| 228 | 335 | 15 | 11 | 150 | 66 | 2 | 10 | 29 | 135 | 2 | 1 | 395 | 3.8 | 42.3 | 44.5 |
| White Bifid × Mini Basic. | |||||||||||||||

| 
| 
| 
| 
| 
| 
| 
| 
|  | ||||||
| 344 | 31 | 109 | 2 | 58 | 41 | 2 | 0 | 6 | 1 | ||||||
SUMMARY OF THE PREVIOUSLY DETERMINED CROSS-OVER VALUES.
SUMMARY OF THE PREVIOUSLY DETERMINED CROSS-OVER VALUES.
The data of the earlier papers, namely, Dexter, 1912; Morgan, 1910c, 1911a, 1911f, 1912f, 1912g; Morgan and Bridges, 1913; Morgan and Cattell, 1912 and 1913; Safir, 1913; Sturtevant, 1913 and 1915; and Tice, 1914, have been summarized in a recent paper by Sturtevant (Sturtevant, 1915) and are given here in table 64. Our summary combines three summaries of Sturtevant, viz, that of single crossing-over and two of double crossing-over.
The data from earlier studies, specifically Dexter, 1912; Morgan, 1910c, 1911a, 1911f, 1912f, 1912g; Morgan and Bridges, 1913; Morgan and Cattell, 1912 and 1913; Safir, 1913; Sturtevant, 1913 and 1915; and Tice, 1914, have been summarized in a recent paper by Sturtevant (Sturtevant, 1915) and are presented here in table 64. Our summary merges three summaries from Sturtevant, which includes one for single crossing-over and two for double crossing-over.
Table 64.—Previously published data summarized from Sturtevant, 1915.
Table 64.—Data that was published earlier, summarized from Sturtevant, 1915.
| Factors. | Total. | Cross-overs. | Cross-over values. |
| Yellow white. | 46,564 | 498 | 1.07 |
| Yellow vermilion. | 10,603 | 3,644 | 33.4 |
| Yellow miniature. | 18,797 | 6,440 | 34.3 |
| Yellow rudimentary. | 2,563 | 1,100 | 42.9 |
| Yellow bar. | 191 | 88 | 46.1 |
| White vermilion. | 15,257 | 4,910 | 32.1 |
| White miniature. | 41,034 | 13,513 | 32.8 |
| White rudimentary. | 5,847 | 2,461 | 42.1 |
| White bar. | 5,151 | 2,267 | 44.0 |
| Vermilion miniature. | 5,329 | 212 | 4.0 |
| Vermilion rudimentary. | 1,554 | 376 | 24.1 |
| Vermilion bar. | 7,514 | 1,895 | 25.2 |
| Miniature rudimentary. | 12,567 | 2,236 | 17.8 |
| Miniature bar. | 3,112 | 636 | 20.4 |
| Rudimentary bar. | 159 | 7 | 4.4 |
SUMMARY OF ALL DATA UPON LINKAGE OF GENS IN CHROMOSOME I.
SUMMARY OF ALL DATA ON LINKAGE OF GENES IN CHROMOSOME I.
In table 65 all data so far secured upon the sex-linked characters are summarized. These data include the experiments previously published in the papers given in the bibliography and the experiments given here. The data from experiments involving three or more loci are calculated separately for each value and included in the totals.
In table 65, all the data collected so far on the sex-linked traits are summarized. This data includes the experiments previously published in the papers listed in the bibliography as well as the experiments presented here. The data from experiments involving three or more loci are calculated separately for each value and included in the totals.
Table 65.—A summary of all linkage data upon chromosome I.
Table 65.—A summary of all linkage data on chromosome I.
BIBLIOGRAPHY.
Bridges, Calvin B.
Calvin B. Bridges
1913. Non-disjunction of the sex-chromosomes of Drosophila. Jour. Exp. Zool., 15, p. 587, Nov. 1913.
1913. Non-disjunction of the sex chromosomes in Drosophila. Journal of Experimental Zoology, 15, p. 587, November 1913.
1914. Direct proof through non-disjunction that the sex-linked gens of Drosophila are borne by the X chromosome. Science, 40, p. 107, July 17, 1914.
1914. Direct evidence through non-disjunction that the sex-linked genes in Drosophila are located on the X chromosome. Science, 40, p. 107, July 17, 1914.
1915. A linkage variation in Drosophila. Jour. Exp. Zool., 19, p. 1. July 1915.
1915. A variation in linkage in Drosophila. Journal of Experimental Zoology, 19, p. 1. July 1915.
1916. Non-disjunction as proof of the chromosome theory of heredity. First instalment, Genetics I, p. 1-52; second instalment, Genetics I, No. 2, 107-164.
1916. Non-disjunction as proof of the chromosome theory of heredity. First part, Genetics I, p. 1-52; second part, Genetics I, No. 2, 107-164.
Chambers, R.
Chambers, R.
1914. Linkage of the factor for bifid wing. Biol. Bull. 27, p. 151, Sept. 1914.
1914. Linking the factor for bifid wing. Biol. Bull. 27, p. 151, Sept. 1914.
Dexter, John S.
Dexter, John S.
1912. On coupling of certain sex-linked characters in Drosophila. Biol. Bull. 23, p. 183, Aug. 1912.
1912. On the combination of specific sex-linked traits in Drosophila. Biol. Bull. 23, p. 183, Aug. 1912.
1914. The analysis of a case of continuous variation in Drosophila by a study of its linkage relations. Am. Nat., 48, p. 712, Dec. 1914.
1914. An analysis of continuous variation in Drosophila by studying its linkage relationships. Am. Nat., 48, p. 712, Dec. 1914.
Duncan, F. N.
Duncan, F. N.
1915. An attempt to produce mutations through hybridization. Am. Nat., 49, p. 575, Sept. 1915.
1915. An attempt to produce mutations through hybridization. Am. Nat., 49, p. 575, Sept. 1915.
Hoge, M. A.
Hoge, M. A.
1915. The influence of temperature on the development of a Mendelian character. Jour. Exp. Zool., 18, p. 241.
1915. The impact of temperature on the development of a Mendelian trait. Journal of Experimental Zoology, 18, p. 241.
Morgan, T. H.
Morgan, T.H.
1910a. Hybridization in a mutating period in Drosophila. Proc. Soc. Exp. Biol. and Med., p. 160, May 18, 1910.
1910a. Hybridization during a changing time in Drosophila. Proc. Soc. Exp. Biol. and Med., p. 160, May 18, 1910.
1910b. Sex-limited inheritance in Drosophila. Science 32, p. 120, July 22, 1910.
1910b. Sex-limited inheritance in Drosophila. Science 32, p. 120, July 22, 1910.
1910c. The method of inheritance of two sex-limited characters in the same animal. Proc. Soc. Exp. Biol. and Med., 8, p. 17.
1910c. How two sex-limited traits are inherited in the same organism. Proc. Soc. Exp. Biol. and Med., 8, p. 17.
1911a. An alteration of the sex-ratio induced by hybridization. Proc. Soc. Exp. Biol. and Med., 8, No. 3.
1911a. A shift in the sex ratio due to hybridization. Proc. Soc. Exp. Biol. and Med., 8, No. 3.
1911b. The origin of nine wing mutations in Drosophila. Science, 33, p. 496, Mar. 31, 1911.
1911b. The origin of nine wing mutations in Drosophila. Science, 33, p. 496, Mar. 31, 1911.
1911c. The origin of five mutations in eye-color in Drosophila, and their mode of inheritance. Science, April 7, 1911, 33, P. 534.
1911c. The origin of five eye color mutations in Drosophila, and their inheritance. Science, April 7, 1911, 33, P. 534.
1911d. A dominant sex-limited character. Proc. Soc. Exp. Biol. and Med., Oct. 1911.
1911d. A dominant sex-limited trait. Proc. Soc. Exp. Biol. and Med., Oct. 1911.
1911e. Random segregation versus coupling in Mendelian inheritance. Science, 34, p. 384, Sept. 22, 1911.
1911e. Random segregation versus coupling in Mendelian inheritance. Science, 34, p. 384, Sept. 22, 1911.
1911f. An attempt to analyze the constitution of the chromosomes on the basis of sex-linked inheritance in Drosophila. Jour. Exp. Zool., 11, p. 365, Nov. 1911.
1911f. An attempt to analyze the structure of chromosomes based on sex-linked traits in Drosophila. Jour. Exp. Zool., 11, p. 365, Nov. 1911.
1912a. Eight factors that show sex-linked inheritance in Drosophila. Science, Mar. 22, 1912.
1912a. Eight factors demonstrating sex-linked inheritance in Drosophila. Science, Mar. 22, 1912.
1912c. Heredity of body-color in Drosophila. Jour. Exp. Zool., 13, p. 27, July 1912.
1912c. The heredity of body color in Drosophila. Jour. Exp. Zool., 13, p. 27, July 1912.
1912d. The masking of a Mendelian result by the influence of the environment. Proc. Soc. Exp. Zool. and Med., 9, p. 73.
1912d. The masking of a Mendelian outcome by environmental factors. Proc. Soc. Exp. Zool. and Med., 9, p. 73.
1912e. The explanation of a new sex-ratio in Drosophila. Science, 36, p. 718, No. 22, 1912.
1912e. Explaining a new sex ratio in Drosophila. Science, 36, p. 718, No. 22, 1912.
1912f. Further experiments with mutations in eye-color of Drosophila. Jour. Acad. Nat. Sci. Phil., Nov. 1912.
1912f. Further experiments on eye color mutations in Drosophila. Jour. Acad. Nat. Sci. Phil., Nov. 1912.
1912g. A modification of the sex-ratio and of other ratios through linkage. Z. f. ind. Abs. u. Veterb. 1912.
1912g. A shift in the sex ratio and other ratios through linkage. Z. f. ind. Abs. u. Veterb. 1912.
1914a. Another case of multiple allelomorphs in Drosophila. Biol. Bull. 26, p. 231, Apr. 1914.
1914a. Another case of multiple alleles in Drosophila. Biol. Bull. 26, p. 231, Apr. 1914.
1914b. Two sex-linked lethal factors in Drosophila and their influence on the sex-ratio. Jour. Exp. Zool., 17, p. 81, July 1914.
1914b. Two sex-linked lethal factors in Drosophila and their influence on the sex ratio. Jour. Exp. Zool., 17, p. 81, July 1914.
1914c. A third sex-linked lethal factor in Drosophila. Jour. Exp. Zool., 17, p. 315, Oct. 1914.
1914c. A third sex-linked lethal factor in Drosophila. Jour. Exp. Zool., 17, p. 315, Oct. 1914.
1914d. Sex-limited and sex-linked inheritance. Am. Nat., 48, P. 577, Oct. 1914.
1914d. Sex-limited and sex-linked inheritance. Am. Nat., 48, P. 577, Oct. 1914.
1915a. The infertility of rudimentary-winged females of Drosophila. Am. Nat., 49, p. 40, Apr. 1915.
1915a. The inability of female Drosophila with underdeveloped wings to reproduce. Am. Nat., 49, p. 40, Apr. 1915.
1915b. The rôle of the environment in the realization of a sex-linked Mendelian character in Drosophila. Am. Nat., 49, p. 385, July 1915.
1915b. The impact of the environment on the expression of a sex-linked Mendelian trait in Drosophila. Am. Nat., 49, p. 385, July 1915.
Morgan, T. H., and C. B. Bridges.
Morgan, T.H., and C.B. Bridges.
1913. Dilution effects and bicolorism in certain eye-colors of Drosophila. Jour. Exp. Zool., 15, p. 429, Nov. 1913.
1913. The impact of dilution and bicolorism on certain eye colors of Drosophila. Journal of Experimental Zoology, 15, p. 429, Nov. 1913.
Morgan, T. H., and Eleth Cattell.
Morgan, T. H., and Eleth Cattell.
1912. Data for the study of sex-linked inheritance in Drosophila. Jour. Exp. Zool., July, 1912.
1912. Information on sex-linked inheritance in Drosophila. Jour. Exp. Zool., July 1912.
1913. Additional data for the study of sex-linked inheritance in Drosophila. Jour. Exp. Zool., Jan. 1913.
1913. Additional information on sex-linked inheritance in Drosophila. Jour. Exp. Zool., Jan. 1913.
Morgan, T. H., and H. Plough.
Morgan, T.H., and H. Plough.
1915. The appearance of known mutations in other mutant stocks. Am. Nat., 49, p. 318, May 1915.
1915. The emergence of recognized mutations in other mutant strains. Am. Nat., 49, p. 318, May 1915.
Morgan, Sturtevant, Muller, and Bridges. The mechanism of Mendelian heredity. Henry Holt & Co., 1915.
Morgan, Sturtevant, Muller, and Bridges. The mechanism of Mendelian heredity. Henry Holt & Co., 1915.
Morgan, T. H., and S. C. Tice.
Morgan, T.H., and S. C. Tice.
1914. The influence of the environment on the size of the expected classes. Biol. Bull., 26, p. 213, Apr. 1914.
1914. The impact of the environment on the expected class sizes. Biol. Bull., 26, p. 213, Apr. 1914.
Rawls, Elizabeth.
Rawls, Elizabeth.
1913. Sex-ratios in Drosophila ampelophila. Biol. Bull. 24, p. 115, Jan. 1913.
1913. Sex ratios in Drosophila ampelophila. Biol. Bull. 24, p. 115, Jan. 1913.
Safir, S. R.
Safir, S. R.
1913. A new eye-color mutation in Drosophila and its mode of inheritance. Biol. Bull. 25, p. 47, June 1913.
1913. A new eye color mutation in Drosophila and its inheritance. Biol. Bull. 25, p. 47, June 1913.
Stark, M. B.
Stark, M.B.
1915. The occurrence of lethal factors in inbred and wild stocks of Drosophila. Jour. Exp. Zool., 19, p. 531-538. Nov. 1915.
1915. The existence of lethal factors in both inbred and wild populations of Drosophila. Journal of Experimental Zoology, 19, pages 531-538. November 1915.
Sturtevant, A. H.
Sturtevant, A.H.
1913. The linear arrangement of six sex-linked factors in Drosophila as shown by their mode of association. Jour. Exp. Zool., Jan. 1913.
1913. The linear arrangement of six sex-linked factors in Drosophila has been shown by their associations. Jour. Exp. Zool., January 1913.
1915. The behavior of the chromosomes as studied through linkage. Z. f. Ind. Abs. u. Vereb. 1915.
1915. Research on chromosome behavior via linkage. Z. f. Ind. Abs. u. Vereb. 1915.
Tice, S. C.
Tice, S. C.
1914. A new sex-linked character in Drosophila. Biol. Bull., Apr., 1914.
1914. A new sex-linked trait in Drosophila. Biological Bulletin, April 1914.
Whiting, P. W.
Whiting, P. W.
1913. Viability and coupling in Drosophila. Am. Nat., 47, p. 508, Aug. 1913.
1913. Viability and coupling in Drosophila. American Naturalist, 47, p. 508, August 1913.
DESCRIPTIONS OF PLATES.
Plate I.
Plate I.
Fig. 1. Normal ♀.
Fig. 1. Regular Female.
Fig. 2. Sable ♀.
Fig. 2. Sable Female.
Fig. 3. Lemon ♂.
Fig. 3. Male Lemon.
Fig. 4. Abnormal abdomen ♀.
Fig. 4. Abnormal abdomen (female).
Fig. 5. Abnormal abdomen ♀.
Fig. 5. Abnormal abdomen (female).
Fig. 6. Yellow ♀.
Fig. 6. Yellow female.
Plate II.
Plate II.
Fig. 7. Eosin, miniature, black ♂.
Fig. 7. Eosin, mini, black ♂.
Fig. 8. Eosin, miniature, black ♀.
Fig. 8. Eosin, mini, black ♀.
Fig. 9. Cherry.
Fig. 9. Cherry.
Fig. 10. Vermilion.
Fig. 10. Vermilion.
Fig. 11. White.
Fig. 11. White.
Fig. 12. Bar (from above).
Fig. 12. Bar (top view).
Fig. 13. Bar (from side).
Fig. 13. Side View of Bar.
Fig. 14. Spot ♀ (abdomen from above).
Fig. 14. Spot female (abdomen viewed from above).
Fig. 15. Spot ♀ (abdomen from side).
Fig. 15. Spot female (abdomen viewed from the side).
Fig. 16. Spot ♂ (abdomen from above).
Fig. 16. Male Spot (abdomen viewed from above).
Fig. 17. Spot ♂ (abdomen from side).
Fig. 17. Male spot (side view of the abdomen).


Notes
[1] For a fuller discussion see "The Mechanism of Mendelian Heredity" by Morgan, Sturtevant, Muller, and Bridges. Henry Holt & Co., 1915.
[1] For a more in-depth discussion, check out "The Mechanism of Mendelian Heredity" by Morgan, Sturtevant, Muller, and Bridges. Henry Holt & Co., 1915.
[2] B. C. here and throughout stands for back-cross.
[2] B. C. here and throughout stands for back-cross.
[3] The first dark body-color mutation "black" (see plate II, figs. 7, 8) had appeared much earlier (Morgan 1911b, 1912c). It is an autosomal character, a member of the second group of linked gens. Still another dark mutant, "ebony," had also appeared, which was found to be a member of the third group of gens.
[4] Wherever reference numbers are given, these denote the pages in the note-books of Bridges upon which the original entries for each culture are to be found.
[4] Wherever reference numbers are provided, these indicate the pages in Bridges' notebooks where the original entries for each culture can be found.
[5] In addition to these expected F1 wild-type females there occurred 13 females of an eye-color like that of the mutant pink. So far as was seen none of the F1 males differed in eye-color from the expected eosin vermilion. Since the eosin vermilion and sable stocks were unrelated and neither was known to contain a "pink" as an impurity, these "pinks" must be due to mutation of an unusual kind. That these "pinks" were really products of the cross is proven by the result of crossing one of them to one of her eosin vermilion brothers, for she showed herself to be heterozygous for eosin, vermilion, and sable.
[5] In addition to the expected F1 wild-type females, there were 13 females with an eye color similar to the mutant pink. So far, none of the F1 males showed any eye color different from the expected eosin vermilion. Since the eosin vermilion and sable stocks were unrelated, and neither was known to have a "pink" impurity, these "pinks" must be the result of an unusual mutation. The fact that these "pinks" were indeed products of the cross is confirmed by the outcome of crossing one of them with one of her eosin vermilion brothers, as she proved to be heterozygous for eosin, vermilion, and sable.
F1 "pink" (Ref. 51 C) ♀ × F1 eosin vermilion ♂.
F1 "pink" (Ref. 51 C) female × F1 eosin vermilion male.
| Reference. | Wild-type. | Eosin vermilion. | Eosin. | Vermilion. | ||||
| ♀ | ♂ | ♀ | ♂ | ♀ | ♂ | ♀ | ♂ | |
| 59 C | 59 | 38 | 43 | 40 | 15 | 9 | 16 | 17 |
In addition to the combinations of eosin and vermilion, sable also appeared in its proper distribution though no counts were made. The four smaller classes are cross-overs between eosin and vermilion. Since no "pinks" appeared the color is recessive, and the brother was not heterozygous for it.
In addition to the combinations of eosin and vermilion, sable also showed up in its usual distribution, although no counts were taken. The four smaller classes are cross-breeds between eosin and vermilion. Since no "pinks" showed up, the color is recessive, and the brother wasn't heterozygous for it.
Two other "pink" females mated to wild males gave similar results in their sons.
Two other "pink" females that mated with wild males produced similar results in their sons.
F1 "pink" ♀ × wild ♂.
F1 "pink" ♀ × wild ♂.
| Reference. | Wild-type ♀. | Wild-type ♂. | Eosin vermilion ♂. | Eosin ♂. | Vermilion ♂. |
| 61 C | 101 | 33 | 37 | 9 | 11 |
These F1 flies should all be heterozygous for "pink." A pair of wild-type flies which were mated gave a 3 : 1 ratio—wild type 51 to "pink" 18. From the "pinks" which appeared in this cross a stock was made which was lost through sterility. Females tested to males of true pink were also sterile, so that no solution can be given of the case.
These F1 flies should all be heterozygous for "pink." A pair of wild-type flies that were mated produced a 3:1 ratio—51 wild-type to 18 "pink." From the "pinks" that resulted from this cross, a stock was established but was lost due to sterility. Females tested with males of true pink were also sterile, so there is no solution to the case.
[6] Purple is an eye-color whose gen is in the second chromosome.
[6] Purple is an eye color determined by a gene on the second chromosome.
[7] The curve published by Miss Stark included by mistake 6 cultures from the succeeding generations, and these coming from only one of the lethals (lethal sb) increase its mode so that the mode of the other lethal (lethal sa) becomes submerged. If these cultures are taken out the curve shows two modes more clearly.
[7] The curve published by Miss Stark mistakenly included 6 cultures from later generations, and these cultures, coming from only one of the lethals (lethal sb), raise its mode, causing the mode of the other lethal (lethal sa) to become overshadowed. If these cultures are removed, the curve displays two modes more distinctly.
[8] The figures to the left in each double column correspond to the symbols above the heavy line, as, in the first example 6,219 white miniature. The similar figure to the right corresponds to the symbol below the heavy line. If no symbols are present below, as in the first example, the column to the right should be read wild-type.
[8] The numbers on the left in each double column match the symbols above the thick line, like in the first example 6,219 white miniature. The corresponding number on the right matches the symbol below the thick line. If there are no symbols below, as in the first example, the column on the right should be interpreted as wild-type.
Download ePUB
If you like this ebook, consider a donation!